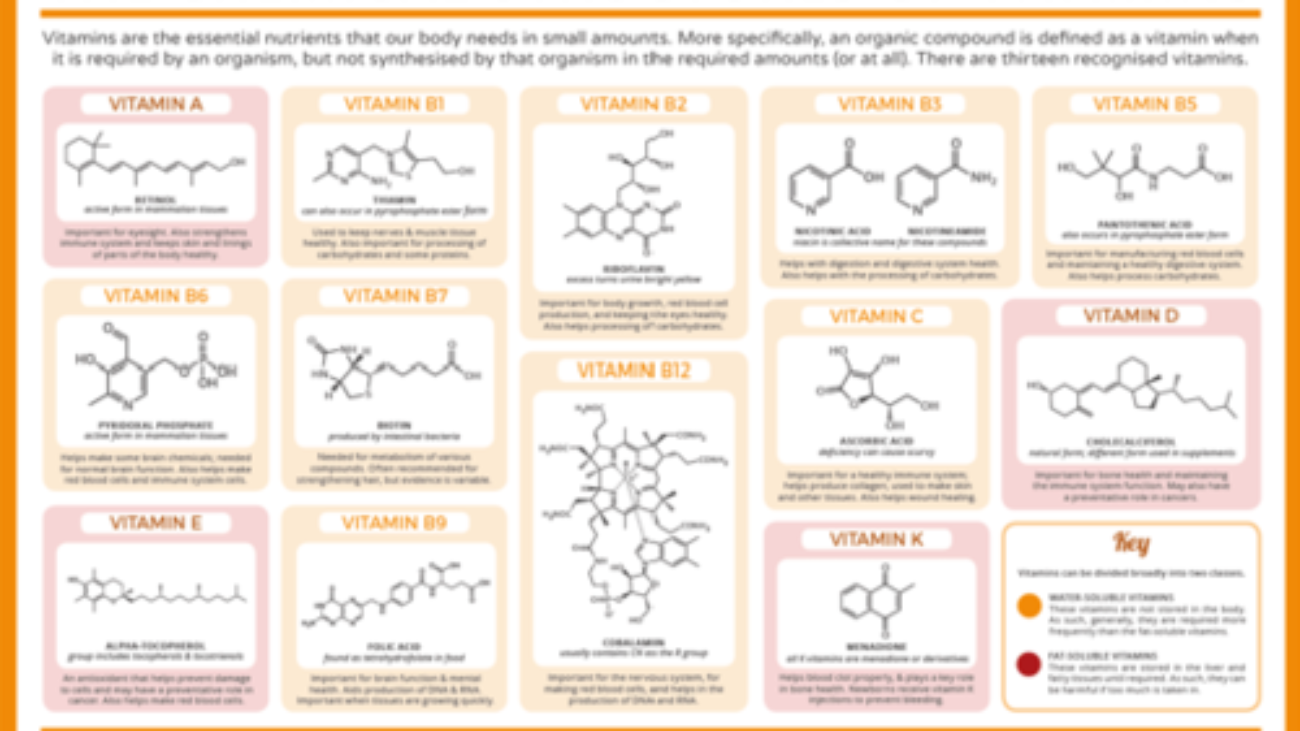
Biochemistry: Vitamins Exam
Test your knowledge with these 30 questions.
Vitamins Exam
Question 1/30
Exam Complete!
Here are your results, .
Your Score
28/30
93%
Test your knowledge with these 30 questions.
Question 1/30
Here are your results, .
Your Score
28/30
93%
Test your knowledge with these 40 questions.
Question 1/40
Here are your results, .
Your Score
38/40
95%
Test your knowledge with these 40 questions.
Question 1/40
Here are your results, .
Your Score
38/40
95%
A 2-year-old boy from Mukono district presents with recurrent episodes of severe bone pain (hands, feet, and sternum pain), jaundice, and fatigue for 3 days.
Laboratory findings reveal:
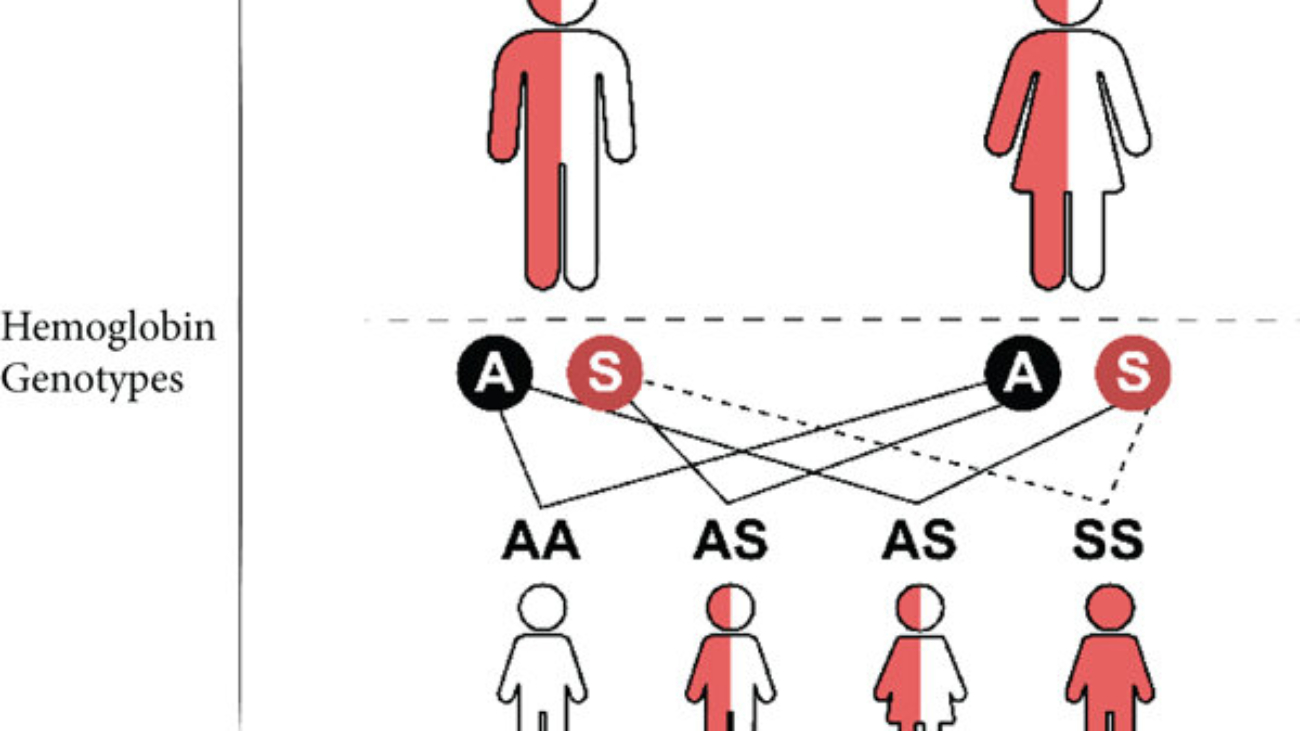
A diagnosis of Vaso-occlusive crisis, and severe anaemia in Sickle Cell Disease was made.
This part requires a detailed breakdown of the specific molecular error in the patient's haemoglobin protein, focusing on the identity of the amino acids and the genetic origin of the mistake.
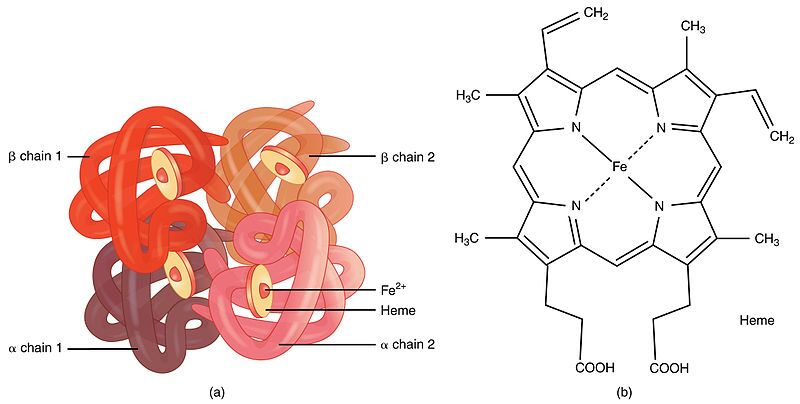
First, it's important to understand what haemoglobin is. Haemoglobin is the primary protein found within red blood cells (erythrocytes) and its main function is to transport oxygen from the lungs to the body's tissues. It is a large, complex protein with a quaternary structure, meaning it is composed of multiple polypeptide subunits. A normal adult haemoglobin molecule (HbA) is a tetramer, consisting of four chains: two identical alpha (α)-globin chains and two identical beta (β)-globin chains. The genetic defect in sickle cell disease specifically affects the gene that provides the instructions for the beta-globin chain.

The defining molecular event in sickle cell disease is a single amino acid substitution at a precise location within the beta-globin polypeptide chain.
In a person with normal adult haemoglobin (HbA), the amino acid at the sixth position from the beginning (the N-terminus) of the beta-globin chain is Glutamic Acid (abbreviated as Glu or E).
In this patient with sickle cell disease, the haemoglobin is abnormal (called HbS). At that exact same sixth position, the Glutamic Acid has been replaced by the amino acid Valine (abbreviated as Val or V).
This single change, Glu6Val, is the sole cause of the disease.

The severity of this substitution is due to the drastically different chemical "personalities" of the R-groups (side chains) of Glutamic Acid and Valine. This position is on the outer surface of the protein, where it is exposed to the watery environment inside the red blood cell.
| Amino Acid | Chemical Class & Properties | Behavior in Water |
|---|---|---|
| Glutamic Acid (Normal) | Its side chain contains a carboxyl group (`-CH₂-CH₂-COOH`). At the neutral pH inside a red blood cell (~7.4), this group loses a proton and becomes negatively charged (`-COO⁻`). Therefore, it is an acidic, polar, and charged amino acid. | Because it is charged and polar, Glutamic Acid is hydrophilic ("water-loving"). It forms favorable interactions with polar water molecules and is perfectly stable on the protein's surface. |
| Valine (Mutant) | Its side chain is an isopropyl group (`-CH(CH₃)₂`), which is a small, branched structure made only of carbon and hydrogen. These bonds are nonpolar. Therefore, Valine is a nonpolar, aliphatic, and neutral amino acid. | Because it is nonpolar, Valine is hydrophobic ("water-fearing"). It is thermodynamically unfavorable for this "oily" side chain to be exposed to water. It will seek to interact with other nonpolar groups to hide from the aqueous environment. |
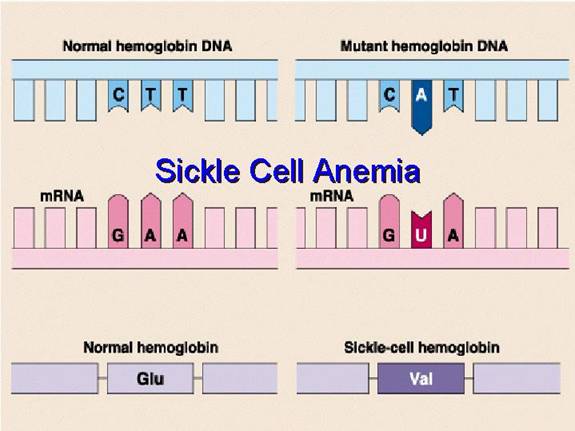
This amino acid error originates from a single change in the DNA sequence of the beta-globin gene. This type of mutation is called a point mutation, specifically a missense mutation because it results in a codon that codes for a different amino acid.
Therefore, a single DNA base change leads to a single mRNA codon change, which in turn leads to the single, catastrophic amino acid substitution that defines sickle cell disease.
This section explains the step-by-step process of how the single Glu6Val substitution causes the haemoglobin to malfunction and leads to the patient's observed symptoms.

The key event is the behavior of HbS when it is in the deoxygenated state. In the oxygenated state (in the lungs), HbS functions almost normally as an oxygen carrier.
Shape Distortion: These long, stiff haemoglobin polymers grow to be longer than the diameter of the red blood cell itself. They physically push against the cell membrane from the inside, distorting the cell from its normal, flexible biconcave disc shape into a rigid, elongated, crescent or "sickle" shape.
Loss of Deformability: This sickling process causes a dramatic loss of the cell's flexibility. It becomes hard and unable to deform. This process is initially reversible if the cell becomes reoxygenated, but repeated sickling events cause permanent membrane damage, leading to irreversibly sickled cells.
The physical properties of these sickled cells are directly responsible for the patient's symptoms:
Knowing that the core problem is a hydrophobic amino acid causing polymerization allows for the design of targeted therapies.
This approach aims to reduce the relative concentration of the problematic HbS.
This is the most direct chemical approach, aiming to stop the Valine from interacting with its target.
This is the most fundamental approach, aiming to fix the DNA instruction so the correct amino acid is made.

Test your knowledge with these 25 questions.
Question 1/25
Here are your results, .
Your Score
23/25
92%
Test your knowledge with these 51 questions.
Question 1/51
Here are your results, .
Your Score
48/51
94%

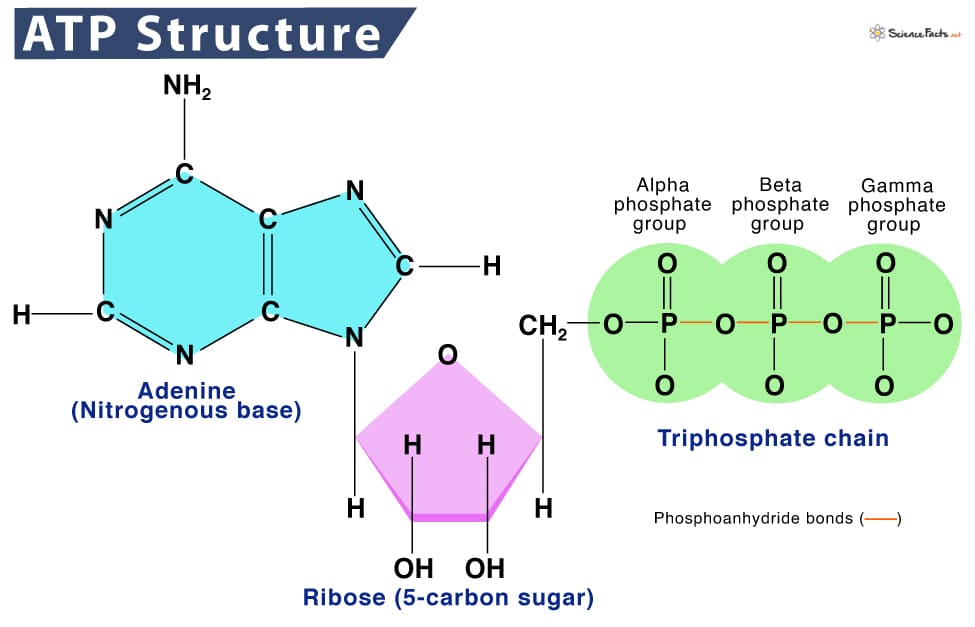
Once the body gets energy from food, it doesn't directly use these complex food molecules to power every single tiny process. Instead, the body converts the chemical energy stored in these food molecules into a much more manageable and readily available form: a special molecule called ATP.
Why is ATP called the "Energy Currency"? Think of it like money. You don't get paid in raw materials; you get paid in money, which you can use to buy whatever you need. Similarly, your body converts energy from diverse food sources into ATP (the "money"). Then, it uses ATP to "pay for" all its energy-requiring processes.
ATP is the direct, usable form of energy for almost all cellular activities.
The key lies in the "high-energy" chemical bonds connecting its three phosphate groups. When your cells need energy, they break off one of the phosphate groups from ATP. This breaking of the bond releases a significant amount of free energy that the cell can immediately use.
ATP → ADP + Pᵢ + Energy
This reaction is reversible. When your body has excess energy, it can use it to reattach the phosphate group to ADP, converting it back into ATP, thus "recharging the battery."
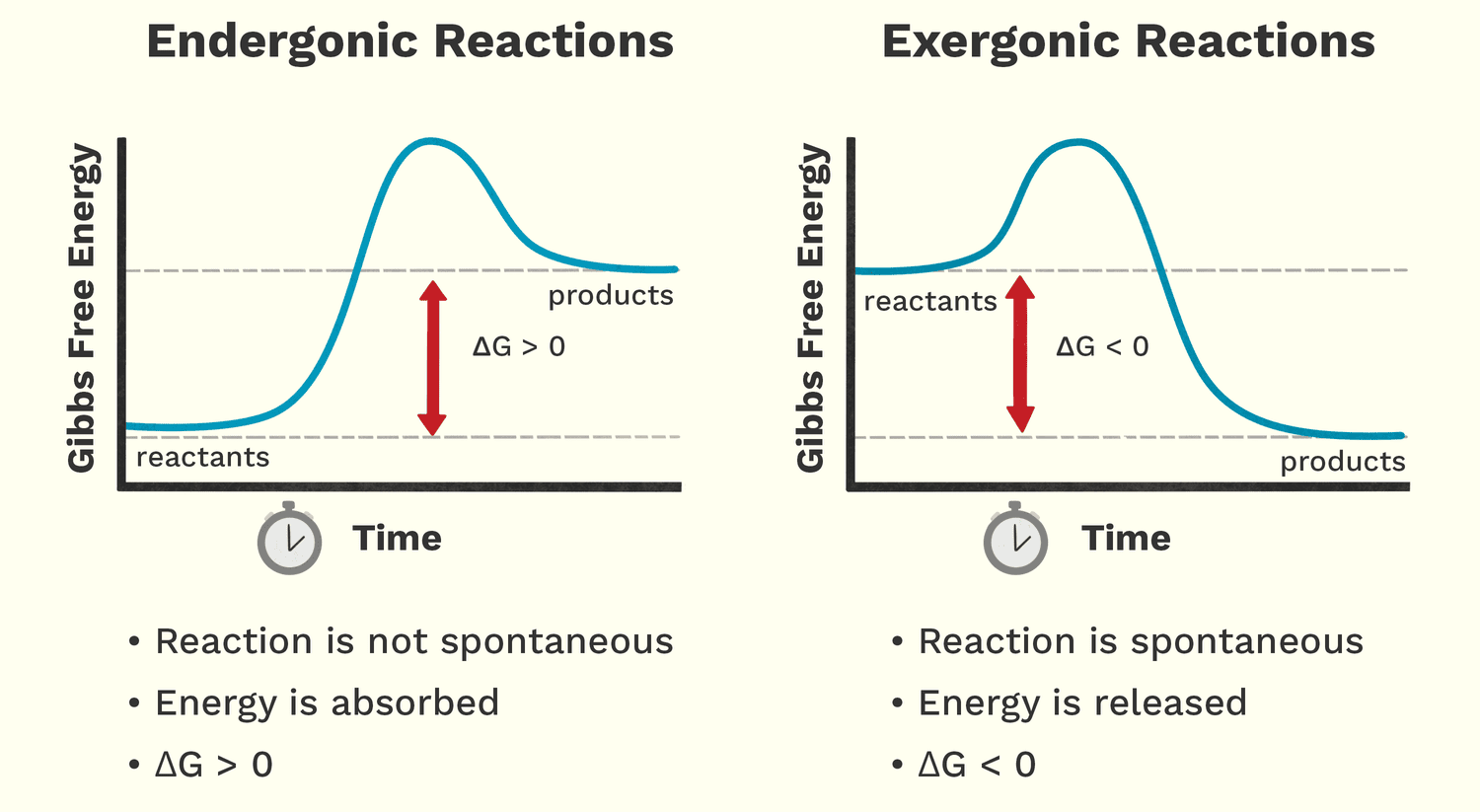
Free energy (Gibbs Free Energy, G) is the amount of energy available to do work within a system. It helps us predict whether a chemical reaction will happen spontaneously (release energy) or require an input of energy.
Analogy: A person moving down a hill is a spontaneous process that releases energy. A person lifting a weight up a hill is a non-spontaneous process that requires energy.
These reactions release free energy and can happen spontaneously. The change in free energy (ΔG) is negative (ΔG < 0).
Biological Examples:
These reactions require an input of free energy and are non-spontaneous. The change in free energy (ΔG) is positive (ΔG > 0).
Biological Examples:
Life thrives by ingeniously linking these two types of reactions together. Cells use the energy released from an exergonic reaction (like ATP breaking down) to drive an endergonic reaction that needs energy. This is called energy coupling. ATP is the perfect intermediate, acting as the bridge that carries energy from energy-releasing pathways to energy-requiring processes.

The overarching scientific field that governs all energy concepts is Thermodynamics. It is a branch of science that deals with the transformation or interconversion of different forms of energy, and how that energy is utilized.
Literally, thermodynamics is about the power of heat or the movement of heat and energy. While "heat" is in the name, it encompasses all forms of energy relevant to biological systems, including light, thermal, chemical, electrical, and mechanical energy.
Thermodynamics is built upon a few fundamental principles known as the Laws of Thermodynamics. These laws are absolute and govern all energy transformations in the universe, including those happening inside the human body.
"Two systems in equilibrium with a third system are in thermal equilibrium with each other."
Meaning: This law defines temperature and is the principle that allows a thermometer to accurately measure a patient's temperature.
Biological Implication: This law underpins the concept of body temperature and thermoregulation. Our bodies constantly strive to maintain a thermal equilibrium (homeostasis).
"Energy cannot be created or destroyed, only transformed from one form to another."
Meaning: The total amount of energy in the universe is constant. You can't get something for nothing.
Biological Implications: Plants don't "make" energy; they transform light energy into chemical energy. When you exercise, you convert chemical energy from food into mechanical energy and heat. Life needs a constant input of energy because organisms are continuously transforming it from external sources to fuel internal processes.
"In any isolated system, the total entropy (disorder) can only increase or remain constant."
Meaning: The universe naturally tends towards a state of greater disorder, randomness, or chaos. Things naturally fall apart; they do not spontaneously become more organized without external effort.
Biological Implications: Living organisms are incredibly complex, highly ordered structures. To maintain this order and fight against entropy, organisms must constantly consume energy. Life is a continuous battle against the Second Law. Every energy transformation results in some energy being "lost" as unusable heat, increasing the entropy of the environment.
"The entropy of a system approaches a constant minimum value as its temperature approaches absolute zero."
Meaning: As a system's temperature gets closer to absolute zero (-273.15 °C), the disorder of the system approaches a minimum. At absolute zero, a perfect crystal would theoretically have zero entropy (perfect order).
Biological Implication: This law highlights the relationship between temperature and molecular motion/disorder. Very low temperatures reduce molecular motion, which is why cryopreservation attempts to halt metabolic processes by drastically reducing temperature and entropy.
Bioenergetics is essentially the application of thermodynamic principles to biological systems. It helps us understand:
These laws have direct clinical applications:
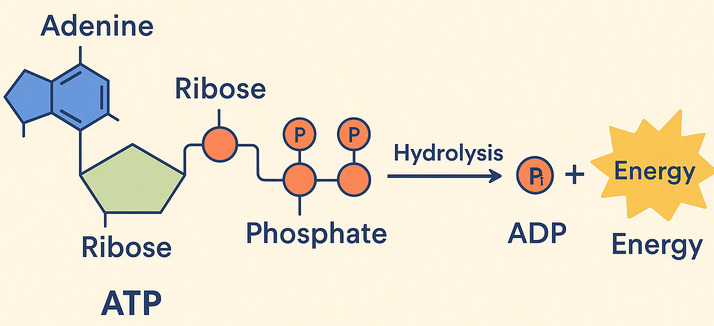
We've already introduced ATP as the energy currency that cells use to "pay for" their work. Now let's understand exactly how this remarkable molecule functions in this essential role.
ATP (Adenosine Triphosphate) is made of adenosine and three phosphate groups. The key to its power lies in the bonds between these phosphate groups, often called "high-energy phosphate bonds."
The term "high-energy" refers to the fact that when these bonds are broken, a significant amount of free energy is readily released. This is because the three negatively charged phosphate groups strongly repel each other, creating strain. Breaking the bond reduces this repulsion, and the remaining molecules (ADP and Pᵢ) settle into a more stable, lower-energy state. The difference in energy is what the cell can harness.
When the cell needs energy, it breaks the outermost phosphate bond in a process called hydrolysis, because a molecule of water (H₂O) is used to break the bond.
ATP + H₂O → ADP + Pᵢ + Free Energy
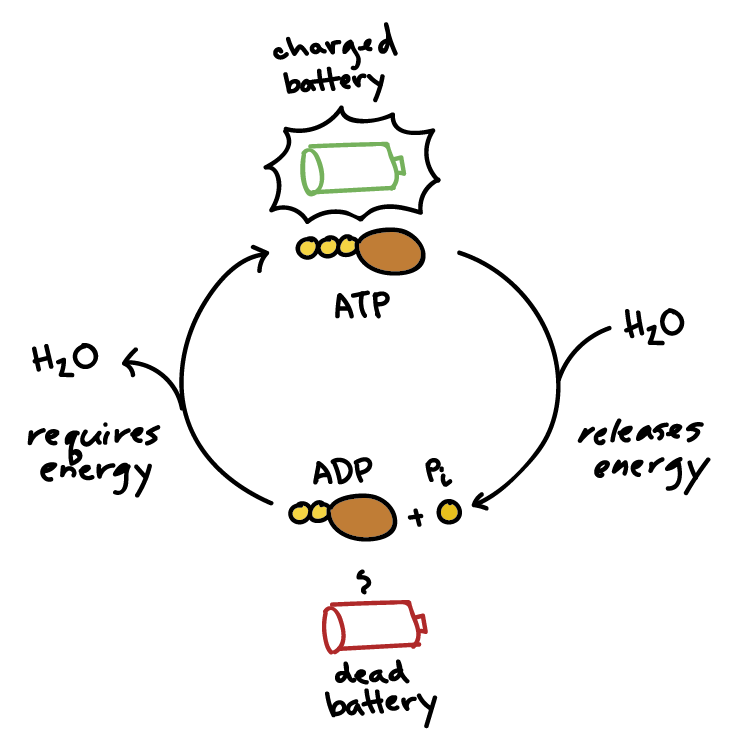
This is truly the magic of ATP! It perfectly acts as the bridge between energy-releasing (exergonic) and energy-requiring (endergonic) processes.
Life depends on a continuous, rapid cycle of ATP breakdown and synthesis:
ADP + Pᵢ + Energy (from food) → ATP + H₂OATP + H₂O → ADP + Pᵢ + Free Energy (for work)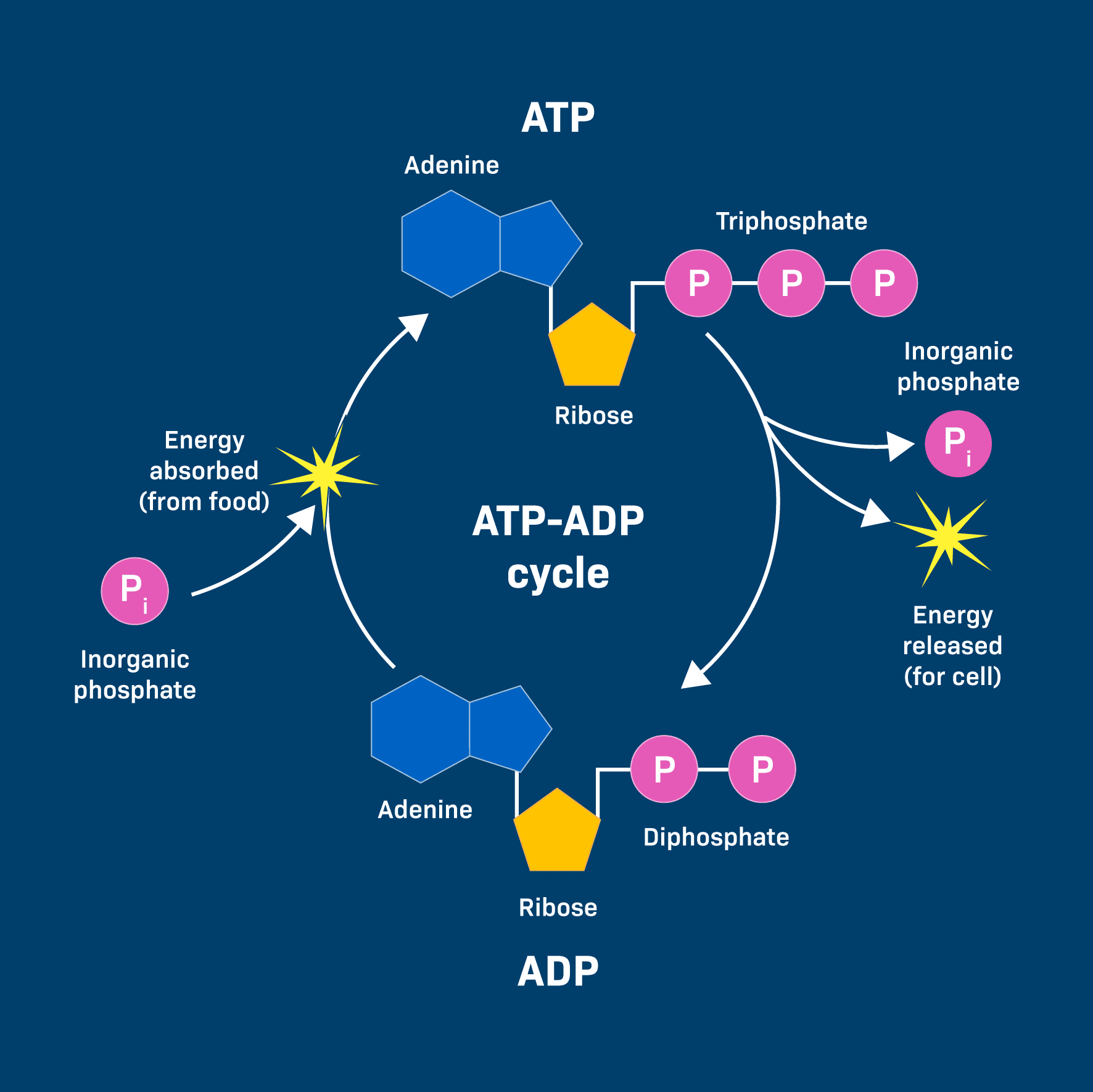
This energy is often transferred through a clever mechanism called phosphorylation. The phosphate group released from ATP is temporarily transferred to another molecule. This energizes the receiving molecule, making it more reactive and priming it to undergo its desired endergonic reaction.
Example: Muscle Contraction
An ATP molecule binds to a muscle protein (myosin). The ATP is hydrolyzed, and the phosphate (Pᵢ) temporarily attaches to the protein (phosphorylation). This causes a change in the protein's shape, leading to the physical contraction (the "work").
So, to summarize the continuous flow of energy that powers life:
Understanding ATP's role is fundamental to comprehending cellular health:

We previously touched upon the Second Law of Thermodynamics, which introduced the powerful idea that things naturally tend towards disorder. This concept is called entropy, and it's a critical component of understanding where "free energy" comes from.
Entropy (S) is a fundamental thermodynamic property that serves as a quantitative measure of randomness or disorder within a system. The more ways particles can be arranged, or the more freely they can move, the higher the entropy.
Analogy: Generally, gases (high entropy, chaotic) have higher entropy than liquids (medium entropy, less ordered), which have higher entropy than solids (low entropy, ordered). Breaking large, complex molecules into smaller, simpler ones also increases entropy.
Enthalpy (H) is essentially the total heat content or the total potential energy contained within a system at constant pressure. We are most interested in the change in enthalpy (ΔH).
This powerful equation is the heart of bioenergetics because it connects these concepts to determine whether a reaction will be spontaneous (exergonic) or require energy (endergonic).
Reactions are most likely to be spontaneous (exergonic) if they release heat (negative ΔH) AND increase disorder (positive ΔS).
6CO₂ + 6H₂O + Light → C₆H₁₂O₆ + 6O₂
C₆H₁₂O₆ + 6O₂ → 6CO₂ + 6H₂O + Energy
We've talked about ATP hydrolysis as releasing energy, but how does that energy actually get used? The primary way is through phosphoryl group transfer, often referred to simply as phosphorylation.
A phosphoryl group transfer is the movement of a phosphate group (Pᵢ) from one molecule to another. ATP is the most common donor. The enzyme-catalyzed transfer of the terminal phosphate group from ATP to a recipient molecule results in a phosphorylated recipient and ADP.
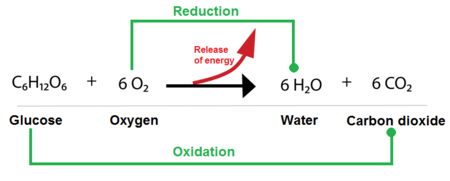
While phosphoryl group transfers are about using energy, oxidation-reduction (redox) reactions are primarily about harvesting and transferring energy from nutrient molecules. This is how cells extract energy from food.
These are always coupled reactions:
A helpful mnemonic is LEO the lion says GER! (Lose Electrons Oxidation, Gain Electrons Reduction).
In biological systems, the transfer of electrons often happens along with the transfer of protons (H⁺), so oxidation often means losing hydrogen atoms (dehydrogenation), and reduction often means gaining them (hydrogenation).
Cells use specialized molecules to pick up and carry electrons. The two most important are:
Test your knowledge with these 20 questions.
Question 1/20
Here are your results, .
Your Score
18/20
90%

At the end of this section, you will be able to understand:
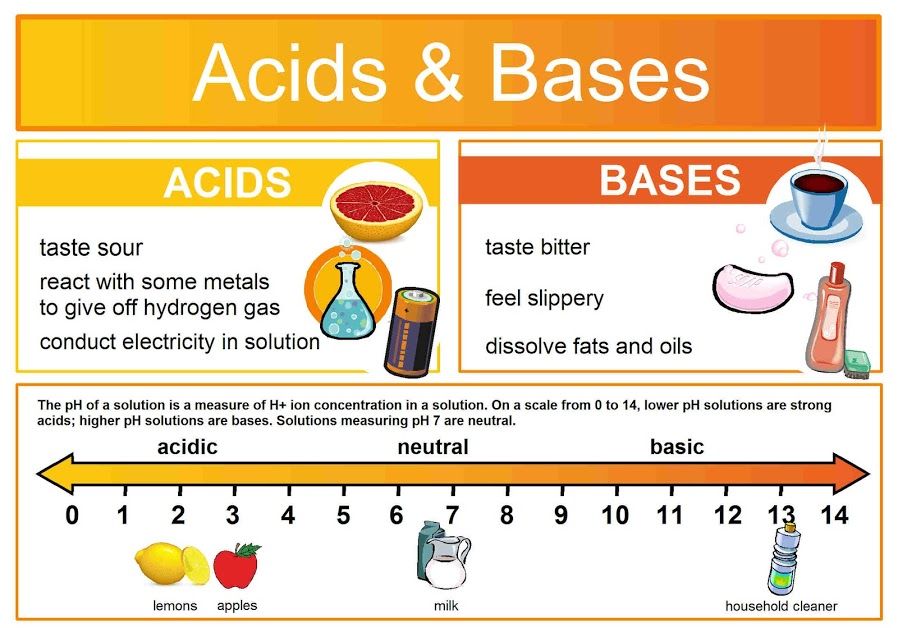
The environment within and around our cells is not static; it's a dynamic chemical soup where countless reactions occur simultaneously. Just like a baker needs to precisely control oven temperature, the "chemical temperature" of our biological systems – its acidity or basicity – must be meticulously maintained within an incredibly narrow range. This control, measured by pH, is paramount for life. Even minor deviations can lead to catastrophic consequences, as the delicate structures of proteins and enzymes are exquisitely sensitive to pH changes. This maintenance of a stable internal pH is a cornerstone of homeostasis.
At the heart of acidity and basicity is one tiny, yet profoundly powerful, particle: the hydrogen ion (H⁺). A hydrogen ion (H⁺) is essentially just a proton. The concentration of these free H⁺ ions in a solution is the ultimate determinant of whether that solution is acidic, neutral, or basic.
An acid is defined as any substance that, when dissolved in an aqueous solution, releases or donates hydrogen ions (H⁺), thereby increasing the concentration of free H⁺ in that solution.
Strength: A strong acid dissociates almost completely in water, releasing nearly all its H⁺ ions. A weak acid only partially dissociates.
Real-World and Physiological Examples:
HCl(aq) → H⁺(aq) + Cl⁻(aq)H₂CO₃(aq) ⇌ H⁺(aq) + HCO₃⁻(aq)A base (or alkali) is any substance that, when dissolved in an aqueous solution, decreases the concentration of H⁺ ions by "accepting" them or by releasing hydroxide ions (OH⁻).
Strength: A strong base dissociates almost completely. A weak base only partially accepts H⁺ or releases OH⁻ ions.
Real-World and Physiological Examples:
NaOH(aq) → Na⁺(aq) + OH⁻(aq)OH⁻(aq) + H⁺(aq) → H₂O(l)HCO₃⁻(aq) + H⁺(aq) ⇌ H₂CO₃(aq)
The definition of acids and bases in this context relies on their behavior in aqueous solutions (where water is the solvent). Water itself can slightly dissociate: H₂O(l) ⇌ H⁺(aq) + OH⁻(aq). In pure water, the concentrations of H⁺ and OH⁻ are equal, making it neutral. Acids disturb this balance by increasing H⁺, and bases disturb it by decreasing H⁺.
Understanding acids and bases is not just theoretical; it's fundamental to clinical practice:
While discussing "hydrogen ion concentration" ([H⁺]) is chemically precise, it's cumbersome. To simplify this, scientists developed the pH scale – a brilliant shorthand that transforms these unwieldy numbers into an easy-to-use linear scale.
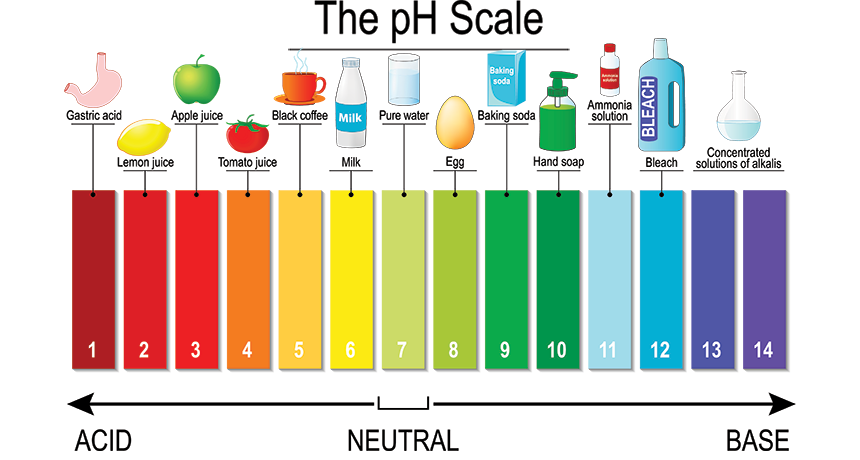
pH literally stands for "potential of Hydrogen" or "power of Hydrogen." It is a numerical scale that quantifies the concentration of hydrogen ions (H⁺) in a solution.
The pH is defined as the negative base-10 logarithm of the hydrogen ion concentration (in moles per liter, M):
pH = −log₁₀[H⁺]
The log₁₀ function makes large numbers manageable, and the negative sign (−) converts the negative results into the positive numbers we see on the scale.
The pH scale typically ranges from 0 to 14.
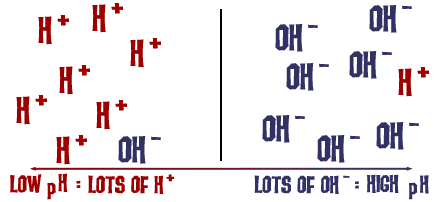
The lower the pH, the higher the [H⁺] concentration.
Examples: Stomach acid (pH 1.5-3.5), lemon juice, coffee.
The concentration of H⁺ equals the concentration of OH⁻.
Examples: Pure water, human tears.
The higher the pH, the lower the [H⁺] concentration.
Examples: Baking soda, ammonia, bleach.
This is perhaps the most important concept about the pH scale. It is logarithmic, not linear. This means that a change of 1 pH unit represents a 10-fold (ten times) change in the actual concentration of H⁺ ions.
Applying the Principle:
Because of this logarithmic nature, even a seemingly small numerical change in pH represents an enormous alteration in the actual concentration of H⁺ ions. This has profound implications for human physiology:
Now that we understand the critical importance of maintaining a stable pH, let's delve into the specific buffer systems that allow the human body to achieve this remarkable feat. These systems are strategically located and exquisitely designed to work in concert, forming a robust defense network.
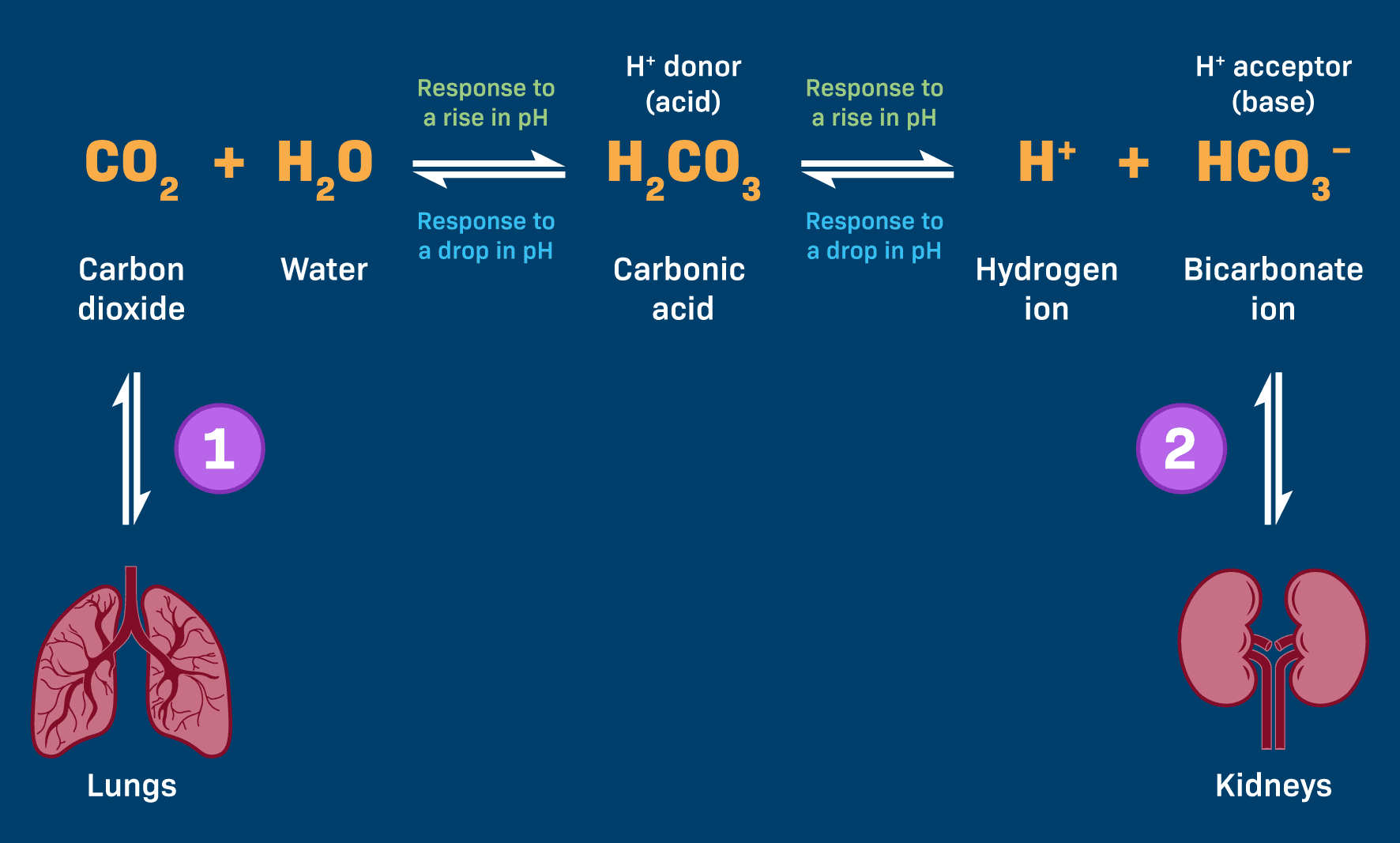
This is arguably the most significant buffer system in the extracellular fluid (ECF). Its power stems from its abundance, the ease with which its components can be regulated, and its intimate connections to both the respiratory (lungs) and renal (kidneys) systems.
These two components exist in a dynamic, reversible equilibrium:
CO₂(g) + H₂O(l) ⇌ H₂CO₃(aq) ⇌ H⁺(aq) + HCO₃⁻(aq)
Every cell produces CO₂ as a waste product. This CO₂ dissolves in blood plasma and, with the help of the enzyme carbonic anhydrase (CA), rapidly reacts with water to form carbonic acid.
CO₂ + H₂O ⇌ H₂CO₃
This step highlights that our body's ongoing metabolic activity directly contributes to the level of the weak acid in this vital buffer system.
Carbonic acid is a weak acid and maintains an equilibrium with its dissociated components: a hydrogen ion (H⁺) and a bicarbonate ion (HCO₃⁻), which is the conjugate weak base ready to accept H⁺ ions.
H₂CO₃ ⇌ H⁺ + HCO₃⁻
This system's elegance lies in its ability to shift the equilibrium in either direction.
The abundant bicarbonate ions (HCO₃⁻) act as proton acceptors, binding to the excess H⁺ to form carbonic acid, a much weaker acid. HCO₃⁻ + H⁺ → H₂CO₃
Respiratory Compensation (Lungs' Role): The carbonic acid formed is unstable and rapidly dissociates back into CO₂ and H₂O. The CO₂ is then exhaled by the lungs. The respiratory system can rapidly increase ventilation (hyperventilation) to "blow off" more CO₂, effectively removing acid from the blood and raising pH.
The carbonic acid (H₂CO₃) component of the buffer dissociates further, releasing more H⁺ ions into the blood to replenish the deficit. H₂CO₃ → H⁺ + HCO₃⁻
Renal Compensation (Kidneys' Role): The kidneys play a slower but more powerful long-term role. They can excrete excess bicarbonate (HCO₃⁻) if the blood is too basic, or reabsorb more bicarbonate if the blood is too acidic. Crucially, they can also excrete H⁺ directly into the urine and generate "new" bicarbonate ions to be returned to the blood.
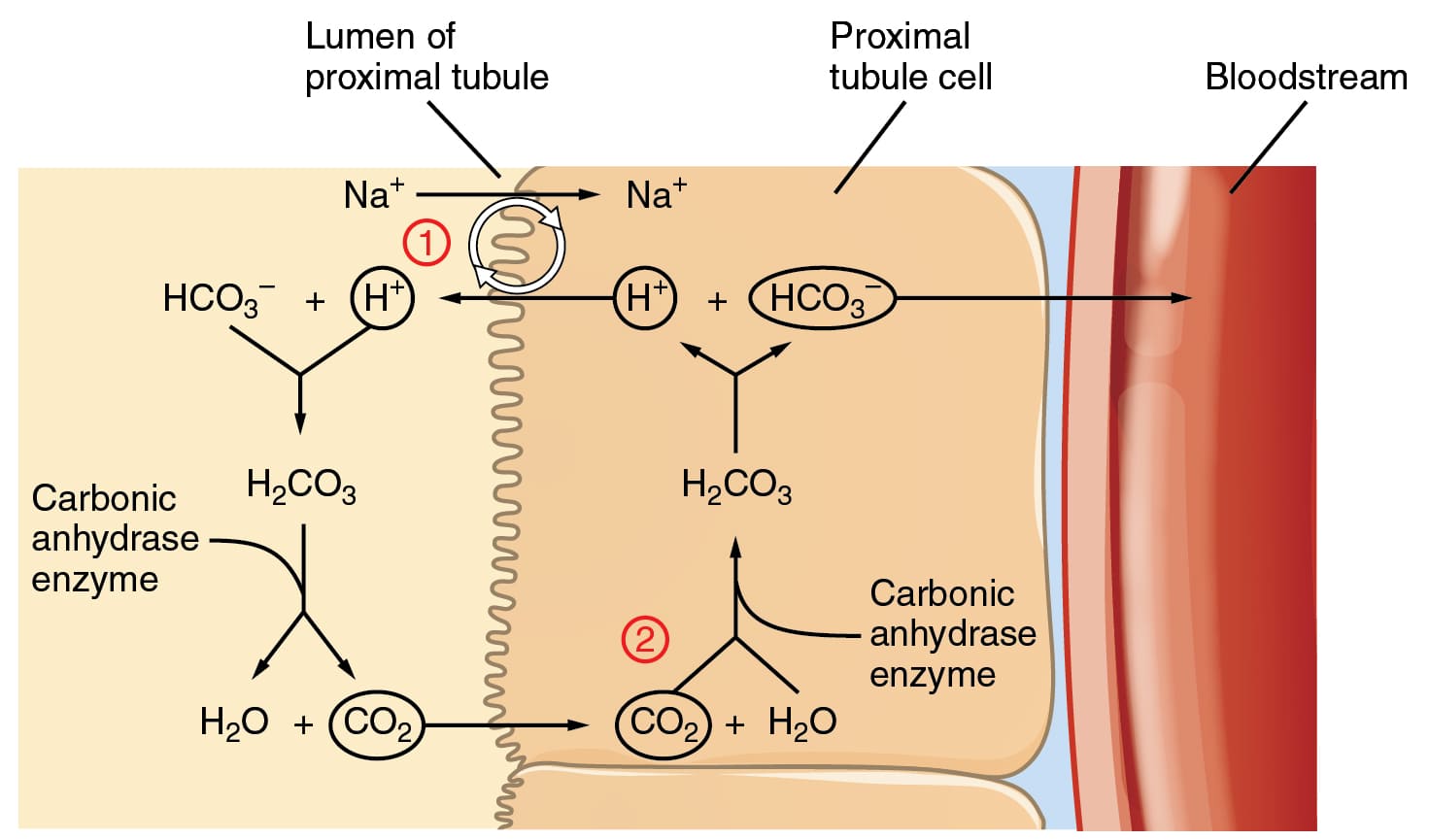
While less quantitatively significant than the bicarbonate system in the blood plasma, the phosphate buffer system plays a vital and specialized role in the intracellular fluid and the urine.
Key Locations: It is a crucial intracellular buffer, where phosphate concentrations are high, protecting enzymes and cellular machinery. It is also very important in urine, allowing the kidneys to excrete a significant amount of acid while keeping urine pH within a manageable range.
The Components:
The Dynamic Equilibrium:
H₂PO₄⁻ ⇌ H⁺ + HPO₄²⁻
How it Works:
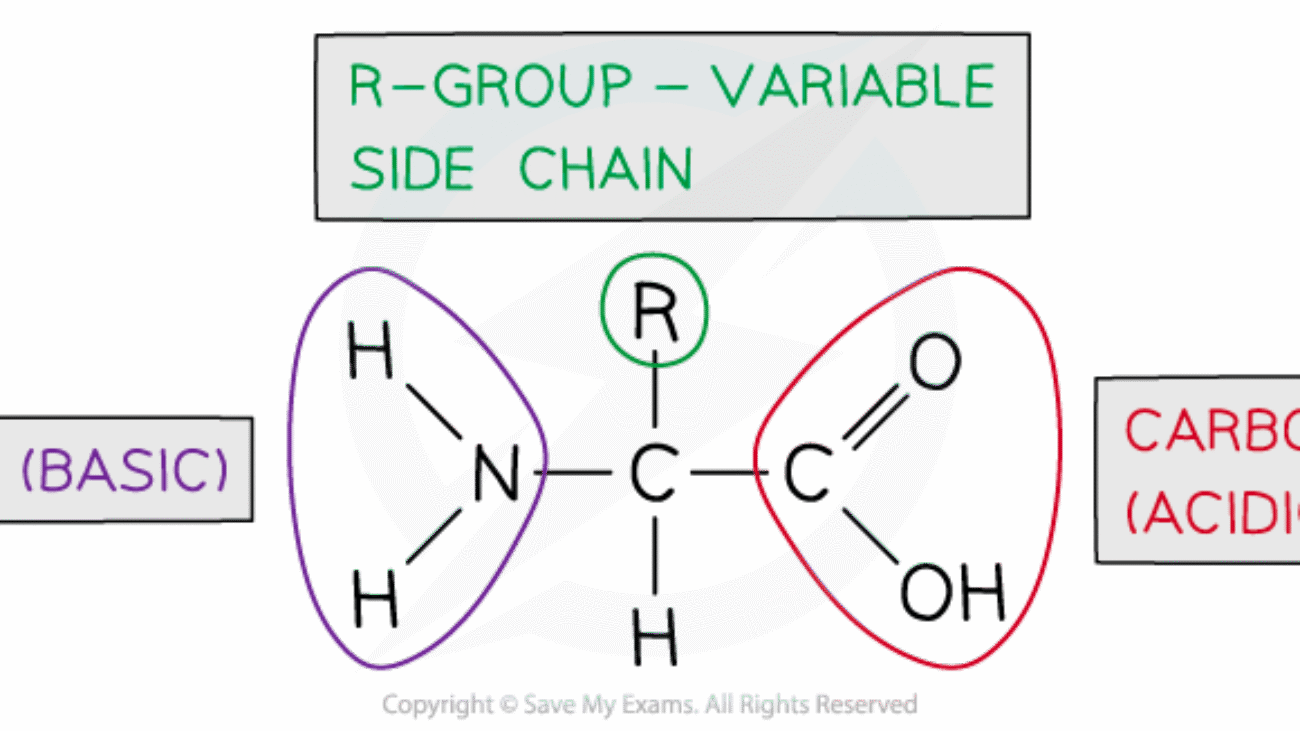
Proteins are the most abundant macromolecules in the body, accounting for approximately 75% of the body's total buffering capacity. Their power comes from their abundance and the unique chemical groups in their amino acid building blocks.
The Components – Amino Acids and Their Buffering Groups:
–NH₂ + H⁺ ⇌ –NH₃⁺–COOH ⇌ –COO⁻ + H⁺A single large protein molecule can contain many of these groups, allowing it to buffer over a wide range of pH values.
Hemoglobin, the protein in red blood cells, is an exceptionally important buffer, especially for CO₂ transport. As CO₂ from tissues enters red blood cells, it is converted to carbonic acid (H₂CO₃), which then dissociates into H⁺ and HCO₃⁻. Hemoglobin immediately binds to these newly generated H⁺ ions.
The Isohydric Shift: Crucially, deoxygenated hemoglobin (found in the tissues) has a greater affinity for H⁺ than oxygenated hemoglobin. This allows it to efficiently buffer the blood in the tissues where acid is being produced. In the lungs, as hemoglobin picks up oxygen, it releases the H⁺, which recombines with HCO₃⁻ to form CO₂ that is then exhaled. This process is vital for preventing a drastic drop in blood pH during CO₂ transport.
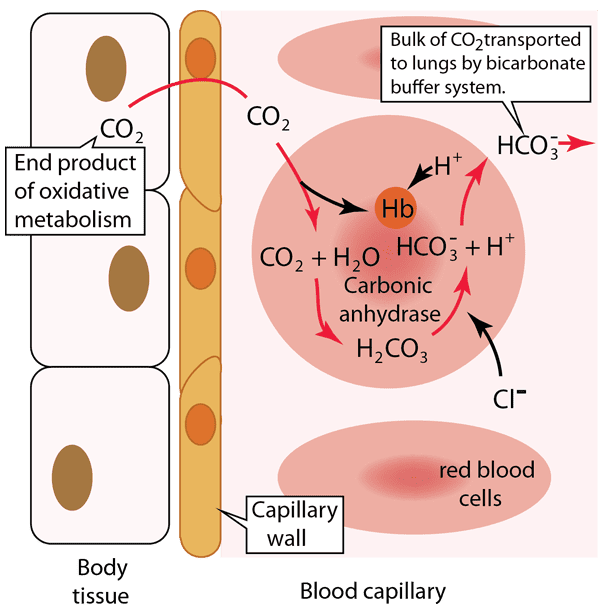
Let's break down the critical process of carbon dioxide transport and pH buffering in the blood, a vital concept for medical students. This diagram illustrates what happens in the body tissues and within a blood capillary, focusing on how CO₂ is handled and how the bicarbonate buffer system, aided by hemoglobin, manages the resulting acid.
Step 1: Carbon Dioxide Production in Body Tissues
Cellular respiration, the process by which cells generate energy, produces carbon dioxide (CO₂) as a waste product. This newly formed CO₂ quickly diffuses out of the body tissue cells because its concentration is higher inside the cells than in the blood. It then crosses the capillary wall to enter the blood.
Step 2: Carbon Dioxide Enters the Red Blood Cell
Once in the blood plasma, a significant portion (about 70-75%) of the CO₂ diffuses into the red blood cells. This is where the magic of the bicarbonate buffer system largely happens for CO₂ transport.
Step 3: Formation of Carbonic Acid and Bicarbonate
Inside the red blood cell, the incoming CO₂ immediately reacts with water (H₂O). This reaction is extremely fast due to the presence of a powerful enzyme called carbonic anhydrase. Carbonic anhydrase rapidly catalyzes the conversion of CO₂ and H₂O into carbonic acid (H₂CO₃), which then quickly dissociates (breaks down) into a hydrogen ion (H⁺) and a bicarbonate ion (HCO₃⁻).
Step 4: Buffering of Hydrogen Ions by Hemoglobin
The hydrogen ions (H⁺) produced are highly acidic. This is where hemoglobin (Hb), the protein responsible for oxygen transport, plays a crucial buffering role. Hemoglobin readily binds to these H⁺ ions, preventing them from accumulating and causing the blood to become too acidic.
Clinical Relevance for Medical: This step is vital because it explains how the body safely handles the acid generated during CO₂ transport without experiencing a severe drop in blood pH (acidosis) at the tissue level.
Step 5: Bicarbonate Ion Transport into Plasma (Chloride Shift)
As bicarbonate ions (HCO₃⁻) accumulate inside the red blood cell, they move out into the blood plasma through a special transporter protein. This is how the bulk of CO₂ is transported to the lungs—in the form of HCO₃⁻ in the plasma. To maintain electrical neutrality, as negatively charged HCO₃⁻ ions move out, negatively charged chloride ions (Cl⁻) move into the red blood cell. This exchange is known as the "chloride shift."
In the body tissues, CO₂ from metabolism enters red blood cells, where it is rapidly converted to H₂CO₃ and then dissociates into H⁺ and HCO₃⁻. Hemoglobin buffers the H⁺, preventing pH changes, while HCO₃⁻ moves into the plasma (via the chloride shift) to be transported to the lungs. This entire process efficiently removes CO₂ from the tissues and minimizes changes in blood pH.
When these red blood cells reach the lungs, the process largely reverses. H⁺ detaches from hemoglobin (as hemoglobin binds oxygen), HCO₃⁻ re-enters the red blood cell, recombines with H⁺ to form H₂CO₃, which then rapidly converts back to CO₂ and H₂O. The CO₂ then diffuses out of the red blood cell and into the alveoli of the lungs to be exhaled.
These buffer systems collaborate in a multi-tiered defense strategy:
The bicarbonate, phosphate, and protein buffer systems provide immediate buffering within milliseconds to seconds. They are always active, chemically neutralizing any H⁺ excess or deficit to "absorb the shock" and buy time for the physiological systems to respond.
The respiratory system acts as a rapid-response physiological buffer, responding within minutes to hours. The lungs can quickly adjust the rate of breathing:
The kidneys are the most powerful and precise regulators of pH, though they act more slowly (hours to days). They are responsible for the long-term maintenance of acid-base balance by:
Test your knowledge with these 20 questions.
Question 1/20
Here are your results, .
Your Score
18/20
90%
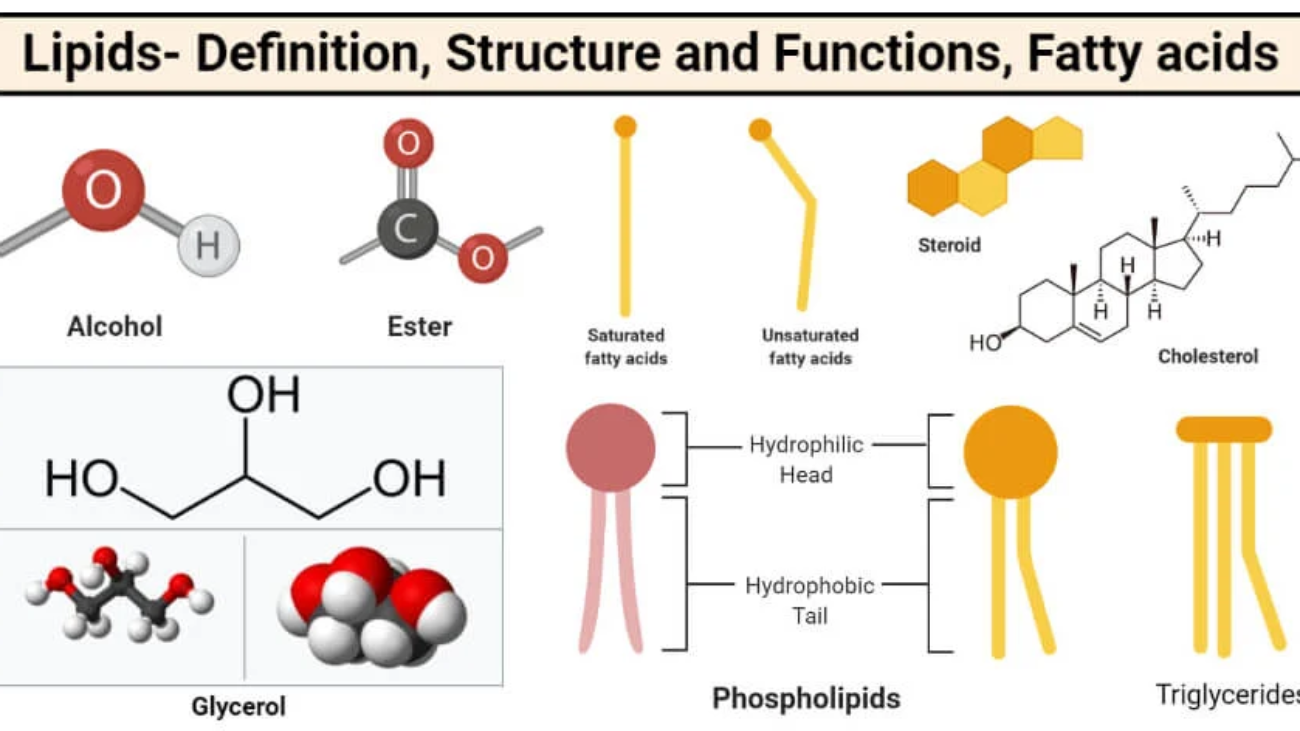
Chemical Bonds:
Water:
We've covered atoms, the fundamental building blocks. However, atoms rarely exist in isolation. The connection that holds atoms together is known as a chemical bond.
Chemical bonds are the powerful attractive forces that hold atoms together to form molecules and compounds. They are entirely about the behavior and interactions of an atom's outermost electrons, called valence electrons. The primary reason atoms form bonds is to achieve a more stable state, typically by having a full outer electron shell.
We've explored atoms, the fundamental building blocks. However, in nature, atoms rarely exist in isolation. They are almost universally connected to other atoms. This crucial connection is known as a chemical bond.
There are two main types of strong chemical bonds that are particularly important in biochemistry and healthcare: Covalent Bonds and Ionic Bonds.
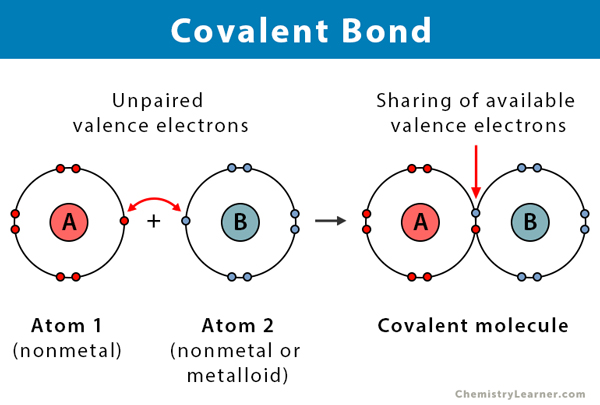
A covalent bond forms when two atoms share one or more pairs of electrons. It's a true partnership, and these are the strong bonds that form the backbone of the vast majority of molecules in our bodies.
Imagine two people who both deeply desire to own a dog, but neither can financially manage the full cost and responsibility alone. They decide to co-own one. They are now "bonded" together by their shared pet. The shared dog represents the shared pair of electrons, and the arrangement benefits both owners. The dog effectively belongs to both, much like the shared electrons are attracted to both atomic nuclei.
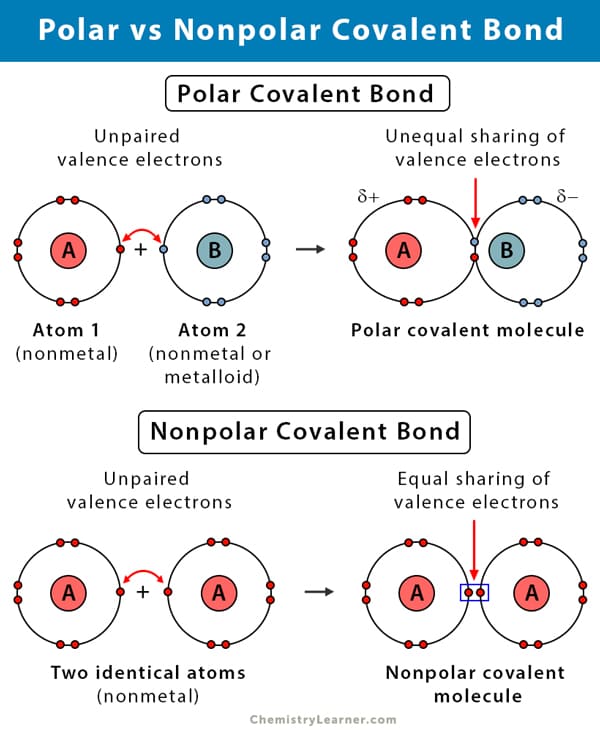
Even within covalent bonds, the sharing of electrons isn't always perfectly equitable. This leads to two important subtypes:

Covalent bonds can also vary in the number of electron pairs shared:
The distinction between nonpolar and polar covalent bonds profoundly impacts biological systems:
The intricate 3D shapes of proteins and DNA are maintained by attractions between the partial positive (δ⁺) and negative (δ⁻) regions of these giant molecules. A disruption in this balance (e.g., from changes in pH or temperature) can cause them to unfold (denature) and lose their function.
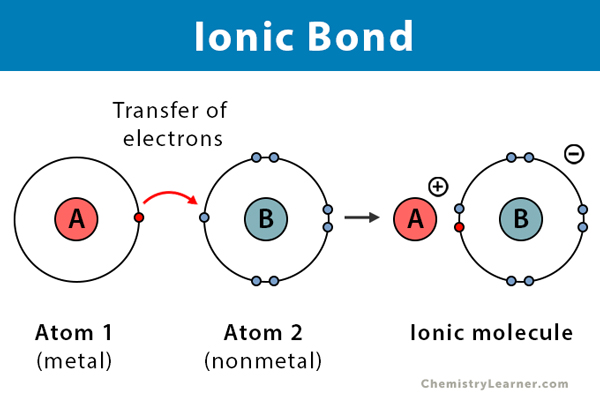
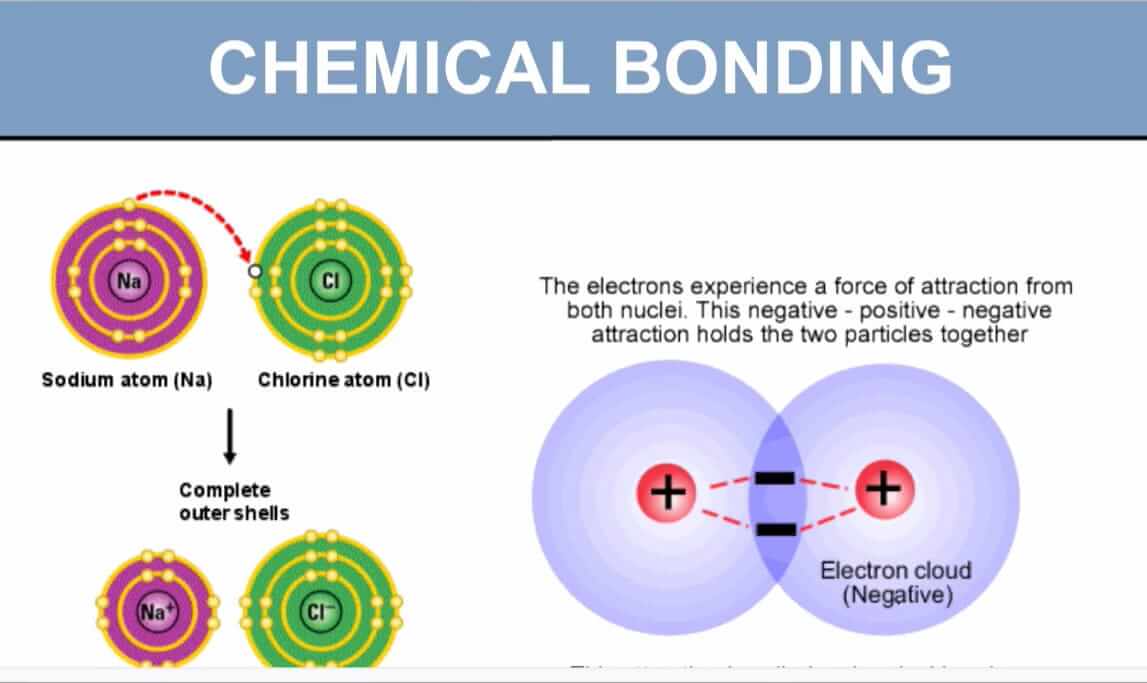
While covalent bonds involve sharing electrons, ionic bonds represent a more dramatic interaction: the complete transfer of one or more electrons from one atom to another.
Instead of co-owning, imagine one person gives their dog to another. The first person feels lighter and happier (like a cation, having lost something). The second person is also happy and stable (like an anion, having gained something). These two are now strongly connected through this exchange.
| Feature | Covalent Bond | Ionic Bond |
|---|---|---|
| Electron Action | Shared between atoms | Transferred from one atom to another |
| Result | Formation of molecules | Formation of ions (cations and anions) |
| Charge | No overall charge (nonpolar) or partial charges (polar) | Full positive and negative charges on ions |
| Strength | Strong | Strong (especially in solid crystals) |
| Key Player | Small difference in electronegativity | Large difference in electronegativity |
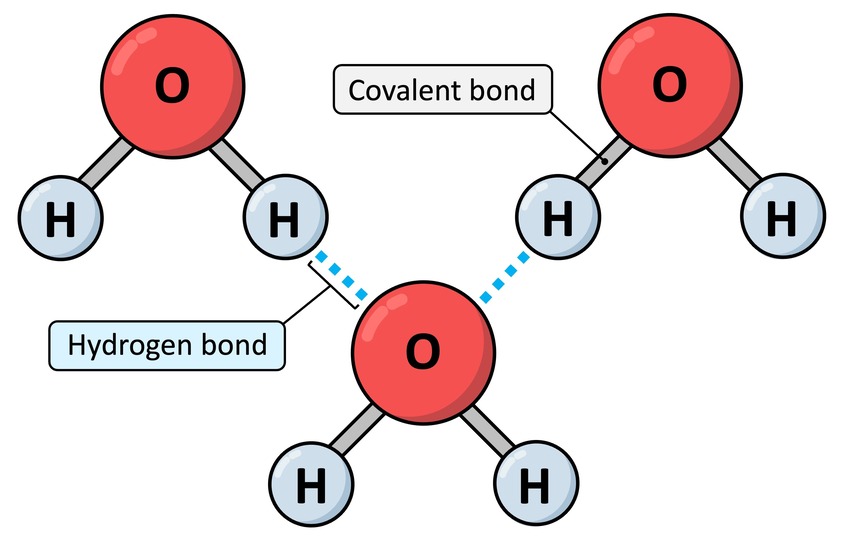
After strong intra-molecular bonds (like covalent and ionic), we now turn to a weaker but incredibly important inter-molecular force: Hydrogen Bonds. These are vital for the structure of water, proteins, and DNA.
Definition: A hydrogen bond is a relatively weak attractive force that forms between a partially positive hydrogen atom (δ+) and a partially negative atom (δ-, usually O or N) on an adjacent molecule or part of a molecule.
While individually weak (5-10% of a covalent bond's strength), the cumulative effect of many hydrogen bonds can create a very significant and stable overall force, like Velcro.
Imagine a person with a very friendly (partially positive) dog (the hydrogen atom). This dog loves to briefly greet other reserved dogs (the partially negative O or N atoms) in the park. Each greeting is a temporary, gentle interaction. But if there are many such friendly dogs, all these brief greetings collectively create a bustling, interconnected social scene.
Hydrogen bonds are the primary reason for many of life's essential chemical properties:
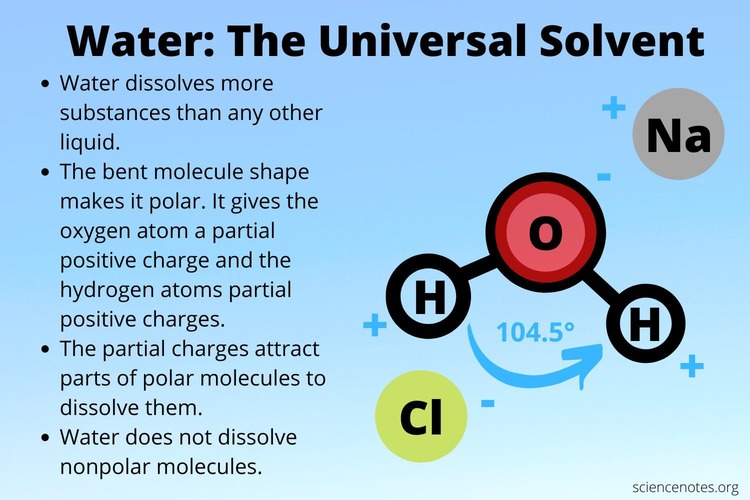
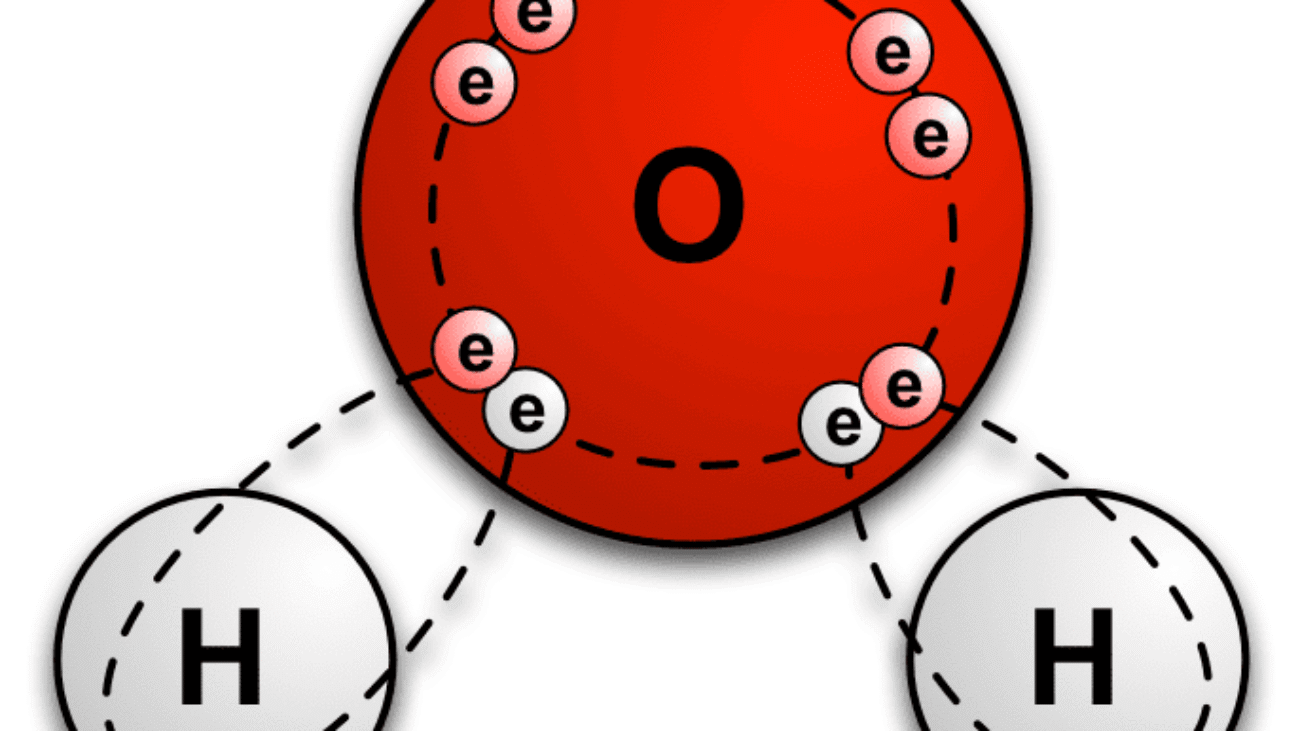
Every single chemical reaction vital for life, from the intricate metabolic pathways that process your last meal to the complex electrochemical signals enabling thought, occurs in an aqueous (water-based) environment. It's no coincidence that water constitutes approximately 60-70% of the human body by weight. Its truly unique and extraordinary properties create the perfect conditions for the chemistry of life to unfold.
The answer to water's exceptional nature lies directly in its molecular structure and, crucially, its unparalleled ability to form hydrogen bonds.
Let's quickly revisit the structure of a water molecule (H₂O):
Because of this separation of charge, a single water molecule acts like a tiny, permanent electrical dipole – essentially, a miniature magnet. This inherent polarity is the foundation for everything that makes water so vital.
When you observe a glass of water, it's not merely a collection of isolated H₂O molecules floating aimlessly. Instead, they are actively and dynamically "sticking" to one another through a continuous network of hydrogen bonds. The slightly positive (δ⁺) Hydrogen of one water molecule is electrostatically attracted to the slightly negative (δ⁻) Oxygen of a neighboring water molecule. Each water molecule can form up to four hydrogen bonds with its neighbors, creating a highly interconnected fluid.
The pervasive network of hydrogen bonds among water molecules gives rise to its extraordinary properties:
Water molecules, being polar, are attracted to other polar or charged surfaces. This is why water "wets" materials like glass and adheres to the internal surfaces of blood vessels.
This is arguably the most important property for biochemistry. Because water molecules are tiny, polar "magnets," they are incredibly adept at pulling apart and surrounding other charged (ionic) or polar molecules.
These are polar or charged molecules that readily dissolve in water.
These are non-polar molecules that do not dissolve in water because they lack charges for water to interact with.
These concepts explain fundamental aspects of human physiology, disease processes, and the efficacy of medications.
Test your knowledge with these 20 questions.
Question 1/20
Here are your results, .
Your Score
18/20
90%
Matter is anything that occupies space and has weight. Literally.
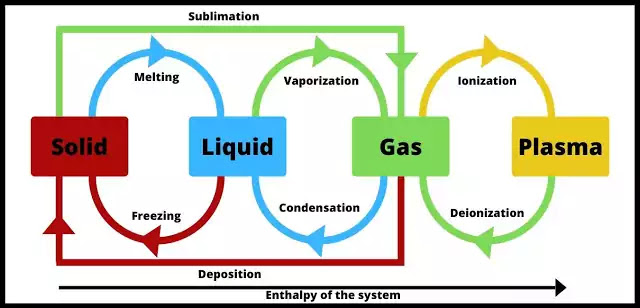
Matter exists in various physical forms, each characterized by distinct particle arrangements and behaviors. Understanding these states is important for comprehending physiological processes and medication properties.
Particles are tightly packed in a fixed, orderly pattern, giving solids a definite shape and volume.
Examples: Bone, pharmaceutical tablets, ice.
Particles are close but can move past each other, allowing liquids to flow and take the shape of their container. They have a definite volume but an indefinite shape.
Examples: Blood, intravenous fluids, water.
Particles are far apart and move randomly and rapidly. They have neither a definite shape nor volume and will expand to fill any container.
Examples: Oxygen, anesthetic agents, air in the lungs.
An ionized gas where some electrons have been stripped from atoms. It is the most abundant state in the universe. It has an indefinite shape and volume but can conduct electricity.
Examples: Lightning, stars.
Objects are distinguished from each other by their physical and chemical properties.
These are characteristics that can be observed or measured without changing the identity of the substance.
Used to differentiate between substances like gold vs. iron, salt vs. sugar, and perfume vs. vinegar.
The mass per unit volume of matter. Materials with a higher density than water sink, while those with lower density float. This is why water isn't used for petrol fires and why helium balloons rise.
The temperature at which a solid changes to a liquid. Cooking pots are made of materials with high melting points.
The temperature at which a liquid changes to a gas. The separation of petroleum oil components is based on their different boiling points.
The resistance of a solid to being scratched or dented. Screwdrivers and building rods are made of extremely hard steel iron.
The ability to allow electricity to flow. Electric wires are made of a conductor (copper) coated in an insulator (plastic).
The ability to allow heat to flow. Cooking pans are made of a good conductor (aluminum) while their handles are made of a bad conductor (wood or plastic).
These properties describe how a substance reacts with other substances to form new materials.
The ability to undergo a chemical reaction. Antacids are used to neutralize stomach acid because their basic properties react with the acid.
The ability to burn or ignite when exposed to heat. Gasoline's high flammability is used to power car engines.
Describes whether a substance is an acid, a base, or neutral. Acidic cleaners are used to dissolve mineral buildup (which is basic).
The ability to damage or destroy another material through a chemical reaction. Bridges and cars are painted to prevent corrosion (rusting).
The degree to which a substance can damage a living organism. Carbon monoxide detectors are installed in homes to protect against poisoning from this toxic gas.
The tendency of a substance to lose electrons, often when combining with oxygen. Antioxidants are added to food to slow down the oxidation that causes spoilage.
The property of an unstable atomic nucleus to spontaneously decay, releasing energy as radiation. In medicine, radiation is used in cancer therapy and medical imaging.
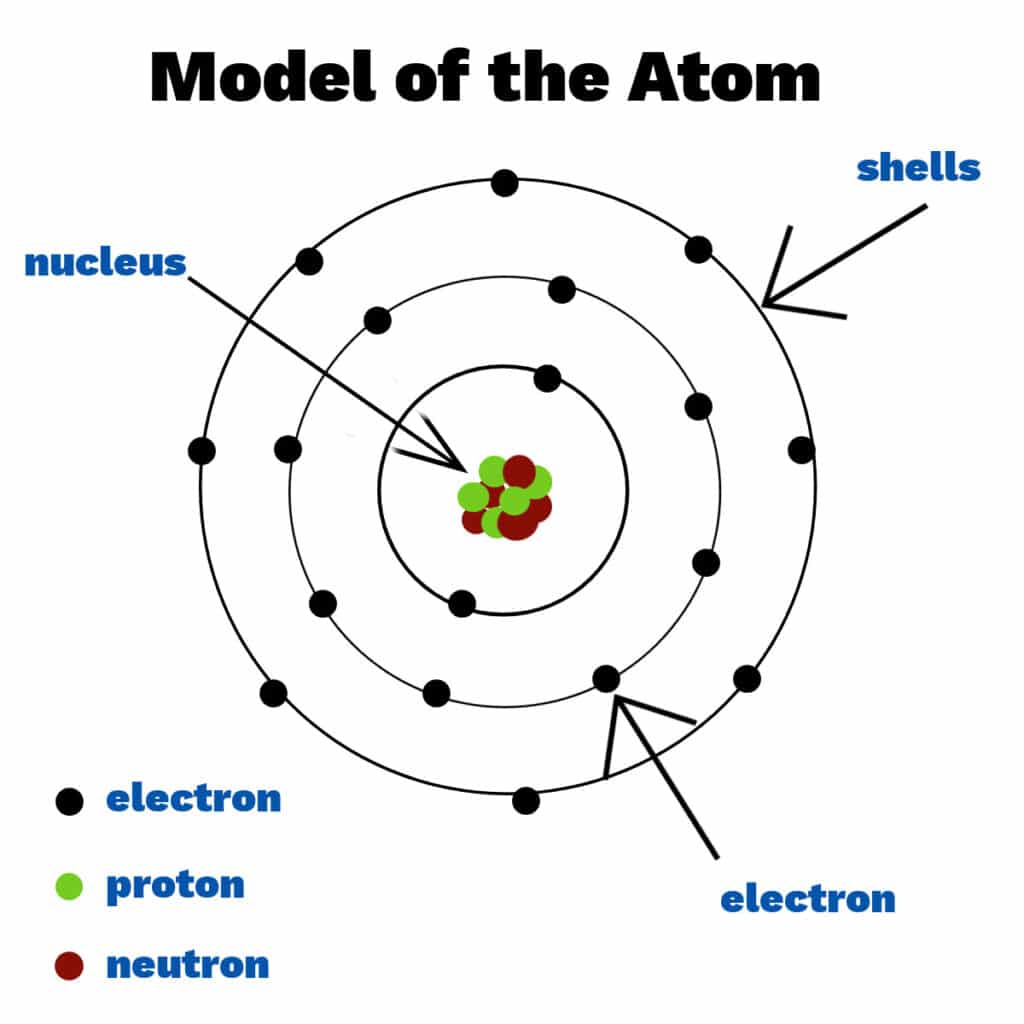
Imagine you have a piece of gold. If you keep cutting it into smaller and smaller pieces, eventually you'd reach the smallest possible piece that still retains the unique characteristics of gold. That irreducible particle is an atom.
An atom is the smallest unit of matter that retains an element's chemical identity.
While it is the smallest chemical unit, it is composed of even smaller, subatomic particles: the electron, proton, and neutron. The central, dense region of an atom is called the nucleus, which holds virtually the entire mass of the atom.
An atom's properties are dictated by the arrangement and characteristics of its subatomic components:
Location: Reside in the atom's central core, the nucleus.
Charge: Possess a positive (+) electrical charge.
Significance: The number of protons (the atomic number) is the defining characteristic of an element. Every carbon atom has 6 protons; changing this number changes the element.
Location: Also found within the nucleus.
Charge: Carry no electrical charge (they are neutral).
Significance: Neutrons stabilize the nucleus. The number of neutrons can vary, creating different isotopes of an element (e.g., Carbon-12 vs. the radioactive Carbon-14).
Location: Orbit the nucleus in specific energy levels or "shells."
Charge: Possess a negative (-) electrical charge.
Significance: Electrons are the primary mediators of chemical bonding between atoms. Their arrangement in the outermost shell dictates an atom's reactivity.
A helpful, though simplified, analogy for atomic structure is a miniature solar system:
While atoms are the fundamental units, matter rarely exists as individual atoms, especially in biological systems.
Examples: An oxygen molecule (O2), a water molecule (H2O), and a glucose molecule (C6H12O6).
In nursing, understanding how atoms form molecules is critical:
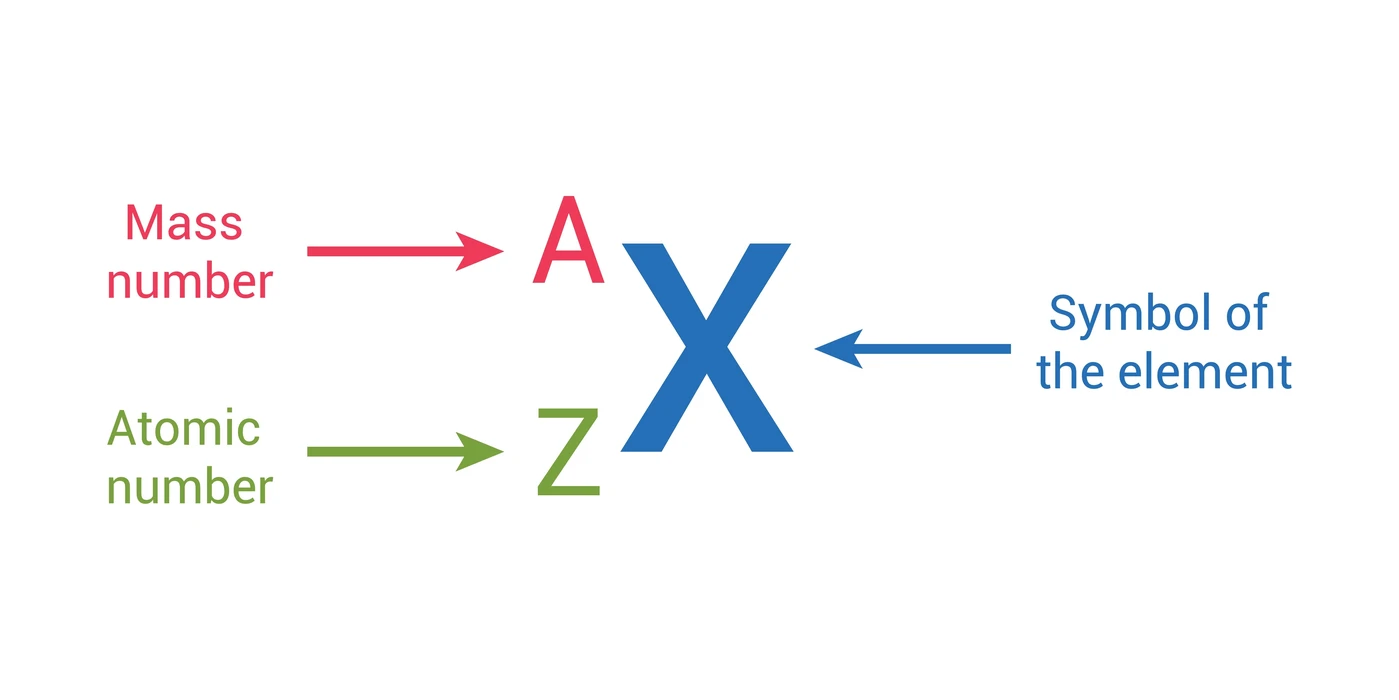
To precisely characterize any atom and understand its behavior, two fundamental numbers are used: the atomic number and the mass number. These concepts are crucial for interpreting chemical formulas, understanding isotopes, and comprehending atomic stability.
Definition: The atomic number (Z) is defined as the exact count of protons residing within an atom's nucleus.
Unique Identifier: This number is the absolute determinant of an element's identity. Each element has a unique atomic number. For example:
Electron Count in Neutral Atoms: For any neutral atom (an atom without an overall electrical charge), the atomic number (number of protons) is precisely equal to the number of electrons.
Definition: The mass number (A) represents the total count of protons and neutrons combined within an atom's nucleus. It essentially provides a measure of the atom's nuclear mass.
Calculation:
Mass Number (A) = Number of Protons + Number of Neutrons
Why Electrons Are Excluded: Electrons are not included because their mass is exceptionally tiny (about 1/1836th of a proton or neutron), making their contribution negligible.
Determining Neutron Count:
Number of Neutrons = Mass Number (A) − Atomic Number (Z)
Consider a common atom of Carbon (C):
While all atoms of a specific element share the same number of protons, they can sometimes differ in their neutron count. This variation gives rise to isotopes.
Definition: Isotopes are atoms of the same element (same number of protons) but with different mass numbers (due to a differing number of neutrons).
Analogy: Think of isotopes as siblings within the same family (the element). They share the same parent (the defining number of protons), but they might have different "weights" due to varying numbers of neutrons.
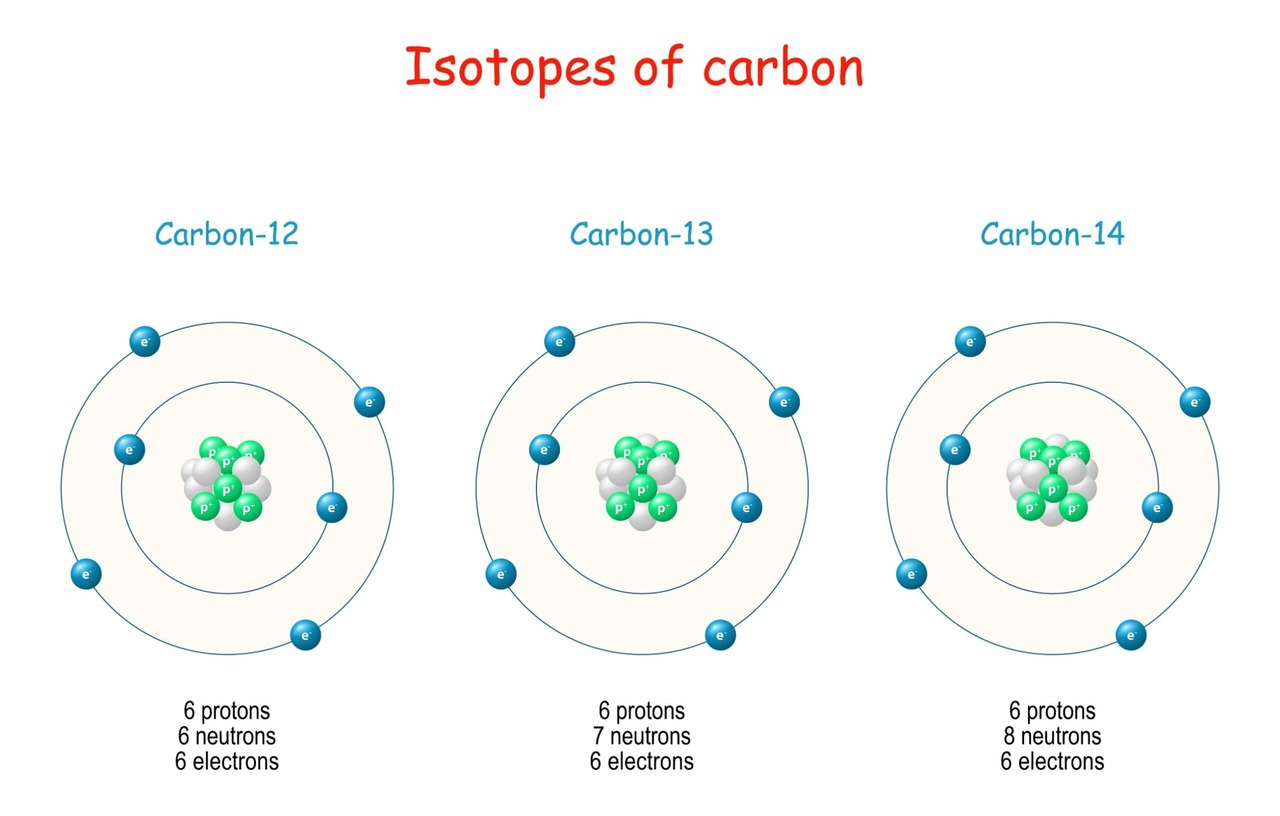
Isotopes are commonly identified by appending their mass number to the element's name.
All carbon atoms have 6 protons.
All oxygen atoms have 8 protons.
Radioactive isotopes (radioisotopes) are invaluable in medicine:
To place isotopes in a broader context, let's briefly revisit the definitions of "element" and "molecule."
An element is a pure substance composed exclusively of atoms that all share the same number of protons (i.e., the same atomic number). Gold, oxygen, hydrogen, and carbon are prime examples.
A molecule is formed when two or more atoms are held together by specific chemical bonds. If a molecule contains atoms from two or more different elements (like H2O), it is also classified as a compound.
When discussing atoms and molecules, their electrical charge is a critical aspect that influences chemical reactivity and biological function. This discussion will temporarily set aside neutrons, as they do not carry an electrical charge.
Definition: An atom is considered neutral when it possesses no net electrical charge. This is achieved because it contains an equal number of protons (positive charge) and electrons (negative charge).
Number of Protons = Number of Electrons
Example (Neutral Carbon): A carbon atom (Atomic Number 6) is neutral when it has 6 protons (+6 charge) and 6 electrons (−6 charge), resulting in a net charge of 0.

When an atom is not neutral, it carries a net electrical charge and is termed an ion. Ions are formed when an atom gains or loses electrons during chemical reactions. The number of protons never changes.
Formation: A cation forms when an atom loses one or more electrons.
Resulting Charge: By losing negative electrons, the atom is left with more protons than electrons, resulting in an overall positive charge.
Example (Sodium Ion, Na+): Neutral Sodium (Na) has 11 protons and 11 electrons. If it loses 1 electron, it has 11 protons (+11) and 10 electrons (−10), for a net charge of +1.
Formation: An anion forms when an atom gains one or more electrons.
Resulting Charge: By gaining negative electrons, the atom has more electrons than protons, resulting in an overall negative charge.
Example (Chloride Ion, Cl−): Neutral Chlorine (Cl) has 17 protons and 17 electrons. If it gains 1 electron, it has 17 protons (+17) and 18 electrons (−18), for a net charge of −1.
The concept of ions is foundational to nearly all biological processes and is critically relevant to nursing practice: