Development of the Central Nervous System
By the end of this highly detailed, exhaustive master guide on neuroembryology, you will be deeply conversant with:
- The molecular inductions and primary germ layer origins orchestrating the Central Nervous System (CNS) formation.
- The complex Sonic Hedgehog (SHH) and Bone Morphogenetic Protein (BMP) signaling pathways governing dorsal-ventral patterning.
- The chronological evolution of primary and secondary brain vesicles and their precise adult derivatives (structures and ventricles).
- The intricate processes of cellular proliferation, radial glial scaffolding, and the Epithelial-to-Mesenchymal Transition (EMT) of the Neural Crest.
- High-yield clinical embryology, including Neural Tube Defects (NTDs), posterior fossa malformations, and migration defects.
SECTION 1: MOLECULAR & EARLY EMBRYONIC INDUCTIONS
The development of the Central Nervous System (CNS) is arguably the most complex, highly orchestrated embryological process in human development. It requires exact spatial and temporal precision, relying heavily on molecular cross-talk between different embryonic tissues.
1.1 Primary Germ Layer Origin & Neurulation
Before the CNS can form, the embryo must undergo Gastrulation (Week 3), converting the bilaminar disc into a trilaminar disc consisting of three primary germ layers: Ectoderm, Mesoderm, and Endoderm. The entire nervous system traces its lineage back to the outermost layer.
- The Origin: The Central Nervous System, Peripheral Nervous System, and the skin epidermis are all derived from the Ectoderm.
- The Induction Process: The ectoderm does not become neural tissue on its own. It requires a chemical command. A specialized rod of mesodermal tissue called the Notochord (the primitive axial skeleton) develops directly beneath the dorsal ectoderm.
- The Notochord's Role: The notochord acts as the primary embryonic organizer. It secretes powerful signaling molecules (like Noggin, Chordin, and Sonic Hedgehog) that diffuse upward into the overlying ectoderm. These signals block inhibitory bone morphogenetic proteins (BMPs), commanding the ectoderm to thicken and transform into the Neural Plate. This specialized tissue is now officially committed to becoming the nervous system (Neuroectoderm).
- Neurulation: The neural plate folds inward to form the neural groove, and the edges (neural folds) fuse together to create the hollow Neural Tube. This closure begins in the cervical (neck) region and zips up towards the head (rostral neuropore closes day 25) and down towards the tail (caudal neuropore closes day 27).
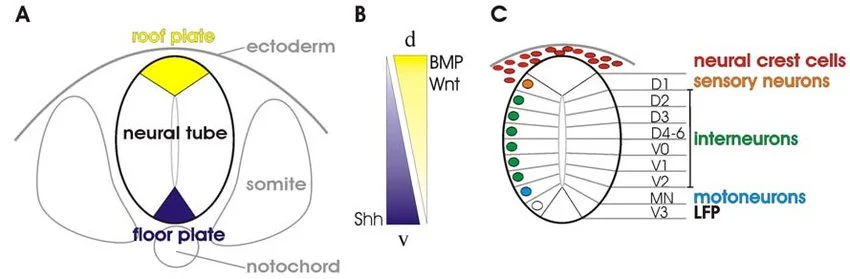
1.2 Sonic Hedgehog (SHH) Signaling Pathway
Sonic Hedgehog (SHH) is one of the most critical morphogens in developmental biology. In the CNS, it is responsible for ventral patterning (making the bottom half of the brain and spinal cord) and the separation of the single primitive brain vesicle into two distinct right and left cerebral hemispheres.
The Complete Signaling Cascade (Step-by-Step):
- Secretion & Gradient: The Notochord secretes SHH protein. The SHH diffuses upward, creating a concentration gradient (highest concentration is ventral, lowest is dorsal). This SHH induces the ventralmost part of the neural tube to become the Floor Plate, which then also begins secreting SHH.
- Receptor Binding: SHH binds to a specific inhibitory cell-surface receptor called Patched-1 (PTCH1).
- Release of Inhibition: Normally, PTCH1 constantly inhibits another transmembrane protein called Smoothened (SMO). When SHH binds PTCH1, the inhibition on SMO is immediately lifted.
- Activation of Transcription: Activated SMO prevents the cleavage of GLI complex proteins. The full, intact GLI zinc-finger transcription factors translocate into the nucleus.
- Gene Expression: GLI activates the expression of specific ventralizing genes, turning the local neural stem cells into motor neurons and interneurons.
Holoprosencephaly (HPE)
Mutations affecting the SHH signaling pathway (e.g., mutations in the SHH gene itself, or maternal exposure to cholesterol synthesis inhibitors, or teratogens like cyclopamine) prevent the embryonic forebrain from properly dividing into two separate hemispheres.
- Result: A spectrum of severe midline defects. The brain remains a single fused lobe (alobar HPE).
- Phenotype: Presents with severe facial abnormalities including cyclopia (a single central eye), proboscis (a tube-like nose above the eye), cleft lip/palate, and a single fused central incisor.
1.3 Dorsal-Ventral Patterning: Alar vs. Basal Plates
As the neural tube develops, it forms a longitudinal groove along its inner lateral walls called the Sulcus Limitans. This acts as a definitive boundary line, splitting the neural tube into a dorsal (top) half and a ventral (bottom) half. This separation dictates the entire sensory and motor organization of the adult CNS.
| Feature | Alar Plate (Dorsal) | Basal Plate (Ventral) |
|---|---|---|
| Location | Dorsal to the Sulcus Limitans. | Ventral to the Sulcus Limitans. |
| Function | SENSORY (Afferent) processing. | MOTOR (Efferent) processing. |
| Molecular Inducer | Induced by BMPs and Wnt secreted by the Roof Plate and overlying surface ectoderm. | Induced by Sonic Hedgehog (SHH) secreted by the Notochord and Floor Plate. |
| Adult Spinal Cord Derivative | Becomes the Dorsal Horn (receives sensory input from dorsal root ganglia). | Becomes the Ventral Horn (contains lower motor neurons projecting to muscles) and Lateral Horn (autonomics). |
| Adult Brainstem Derivative | Migrates laterally. Forms the sensory cranial nerve nuclei (e.g., Trigeminal sensory, Vestibulocochlear). | Remains medially located. Forms the motor cranial nerve nuclei (e.g., Oculomotor, Hypoglossal). |
Sensory = Afferent, Motor = Efferent.
Dorsal = Afferent, Ventral = Efferent.
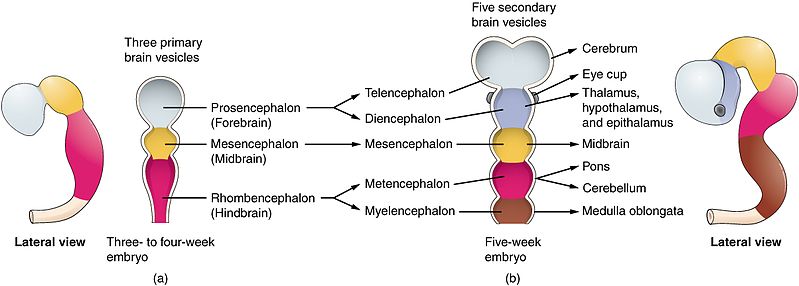
SECTION 2: PRIMARY AND SECONDARY VESICULATION
Once the neural tube forms, its cranial (head) end undergoes a rapid series of swellings and expansions to form the complex structures of the brain. This is known as vesiculation, occurring in two distinct chronological phases.
2.1 The Chronological Evolution of the Neural Tube
Week 4: The 3 Primary Vesicles
At approximately day 28 of gestation, the cranial end of the neural tube expands to form three distinct swellings (primary vesicles):
- Prosencephalon (Forebrain): The most rostral portion.
- Mesencephalon (Midbrain): The middle portion.
- Rhombencephalon (Hindbrain): The caudal portion, continuous with the spinal cord.
Week 5: The 5 Secondary Vesicles
By day 35, the primary vesicles undergo further division. The Prosencephalon and Rhombencephalon split, while the Mesencephalon remains a single vesicle. This creates five secondary vesicles that give rise to the definitive adult brain structures.
- Telencephalon: Rapidly outgrows all other structures, folding over them. It becomes the Cerebral Hemispheres (Cortex, White Matter, Basal Ganglia, Hippocampus, Amygdala, and Olfactory bulbs).
- Diencephalon: The central core. It becomes the Thalamus, Hypothalamus, Epithalamus (Pineal gland), and Subthalamus. Crucially, it also out-pockets to form the optic vesicle, which becomes the Retina and Optic Nerve (CN II).
- Mesencephalon: Does not divide. It simply matures into the adult Midbrain (contains the superior and inferior colliculi, cerebral peduncles, and motor nuclei for CN III and IV).
- Metencephalon: The upper half of the hindbrain. It forms the Pons (ventrally) and the Cerebellum (dorsally).
- Myelencephalon: The lower half of the hindbrain. It forms the Medulla Oblongata, which transitions into the spinal cord.
2.2 Ventricle Mappings
The original hollow cavity running through the center of the embryonic neural tube does not disappear. As the walls of the neural tube thicken into brain tissue, the hollow center expands and contorts into the fluid-filled Ventricular System of the adult brain. Cerebrospinal Fluid (CSF) flows through these connected spaces.
- Telencephalon → Lateral Ventricles: The massive expansion of the cerebral hemispheres pulls the central cavity into two large, C-shaped lateral ventricles (one in each hemisphere). This represents the largest volume of CSF storage.
- Diencephalon → Third Ventricle: The narrow, slit-like space sitting exactly in the midline between the two thalami.
- Mesencephalon → Cerebral Aqueduct (of Sylvius): The cavity drastically narrows in the midbrain. This is the most common site of obstruction, leading to congenital non-communicating hydrocephalus.
- Met/Myelencephalon → Fourth Ventricle: A tent-shaped cavity located posterior to the Pons/Medulla and anterior to the Cerebellum. It features outflow foramina (Luschka and Magendie) that allow CSF to escape into the subarachnoid space.
2.3 The Three Embryonic Brain Flexures
The developing brain tube grows so rapidly that the embryonic skull cannot contain it in a straight line. To fit, the tube must fold upon itself at specific hinges called flexures.
- Cephalic (Mesencephalic) Flexure: Located in the midbrain. This is the only flexure that persists in the adult. It is the reason the human brain is bent at a 90-degree angle relative to the vertical spinal cord.
- Cervical Flexure: Located at the junction between the hindbrain (myelencephalon) and the spinal cord. It straightens out and disappears later in development.
- Pontine Flexure: A unique dorsal fold that occurs between the metencephalon and myelencephalon. As it folds backward, it physically spreads the walls of the neural tube apart like opening a book. This causes the sensory alar plates to shift laterally and the motor basal plates to remain medially—which is why in the brainstem, sensory cranial nerve nuclei are lateral, and motor nuclei are medial.
SECTION 3: CELLULAR PROLIFERATION, MIGRATION, & HISTOGENESIS
The brain requires billions of highly specialized cells, all navigating to exact geometric coordinates in 3D space to wire up functional circuits.
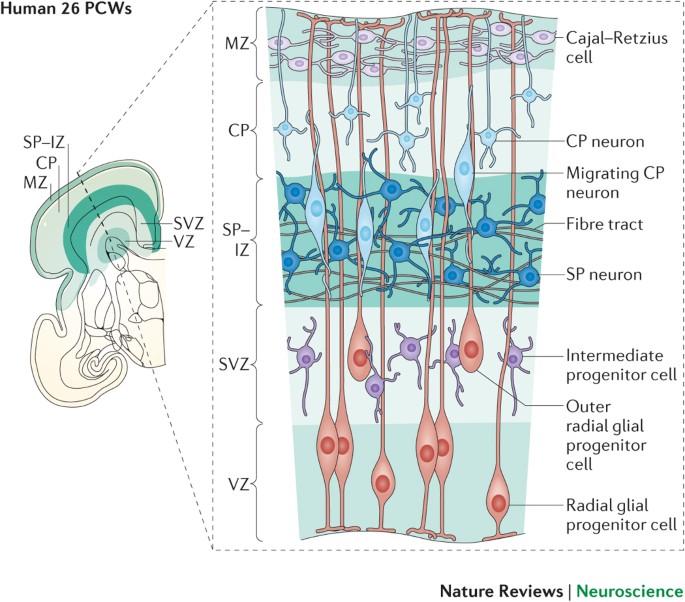
3.1 Radial Glial Cells: The Master Architects
The wall of the developing neural tube is extremely thick. Neuroblasts (baby neurons) are born deep on the inner surface (near the ventricle) and must travel immense distances outward to form the cerebral cortex.
- The Scaffolding: Radial glial cells possess cell bodies in the ventricular zone and extend a single, long, taut physical process all the way to the outer pial surface of the brain.
- The Climb: Newly born neurons physically wrap around these radial glial fibers and crawl up them like a ladder to reach their final destination. This is known as radial migration.
- Inside-Out Development: The cerebral cortex is built in an "inside-out" pattern. The first migrating neurons stop early to form the deepest layer (Layer VI). Subsequent waves of neurons crawl past the older ones to form more superficial layers (Layers V, IV, III, II, and I).
- Molecular Guidance: This migration requires specific chemical signaling. The protein Reelin tells neurons when to detach from the radial glia and stop migrating. The protein Doublecortin (DCX) stabilizes the cytoskeleton during the crawl.
3.2 Dual Cell Origins: The Critical Distinction
In pathology and neuro-oncology, it is mandatory to know the exact embryological origin of every cell type in the brain.
| Embryological Origin | Derived Cell Types | Function / Characteristics |
|---|---|---|
| NEUROECTODERM (From the Neural Tube / Neural Crest) |
Neurons | The excitable signaling cells of the brain. Incapable of division after birth. |
| Astrocytes | Star-shaped support cells. Maintain the Blood-Brain Barrier (BBB), buffer extracellular K+, and form scar tissue (gliosis) after injury. | |
| Oligodendrocytes | Myelinate axons specifically within the Central Nervous System (CNS). One oligodendrocyte can myelinate up to 50 axons. (Targeted in Multiple Sclerosis). | |
| Ependymal Cells | Ciliated cells lining the ventricles. Produce and circulate Cerebrospinal Fluid (CSF). | |
| MESODERM (Invaders from the blood) |
Microglia | The resident macrophages of the CNS. They originate from the embryonic yolk sac (mesoderm), enter the brain through the bloodstream early in development, and act as the brain's innate immune system, phagocytosing debris. |
CRITICAL EXAM POINT: Microglia are the ONLY cells in the brain parenchyma that do NOT come from the ectoderm. They are mesodermal in origin.
3.3 Three Embryonic Layers of the Neural Tube
As the neuroepithelial cells divide, the wall of the neural tube physically thickens and differentiates into three distinct concentric layers/zones:
- Ventricular Zone (Inner layer): The innermost layer immediately lining the central canal/ventricle. This is the primary germinal matrix where intense mitosis and cellular division occurs. After neurogenesis is complete, the remaining cells here mature into the ciliated ependymal cells that line the ventricles.
- Mantle Zone (Intermediate layer): Formed by neuroblasts that have stopped dividing and migrated outward from the ventricular zone. Because it contains the cell bodies of neurons, this layer becomes the Gray Matter of the CNS (e.g., the butterfly-shaped horn of the spinal cord).
- Marginal Zone (Outer layer): The outermost perimeter. It contains almost no cell bodies. Instead, it is composed of the long, myelinated axonal processes sprouting from the neurons in the mantle zone. Because myelin is highly lipid-rich (fatty and white), this layer becomes the White Matter of the CNS (e.g., the outer tracts of the spinal cord).
SECTION 4: NEURAL CREST DIFFERENTIATION & PERIPHERAL STRUCTURES
Often referred to as the "4th Germ Layer," the neural crest is a transient, highly migratory, and incredibly versatile population of cells. Without the neural crest, the Peripheral Nervous System (PNS) and the intricate structures of the face would not exist.
4.1 Epithelial-to-Mesenchymal Transition (EMT)
During neurulation, as the neural folds elevate and prepare to fuse into a tube, the cells right at the apex (the "crest" of the folds) undergo a radical transformation. They lose their tight, cell-to-cell epithelial adhesions by downregulating E-cadherin. They convert into loose, highly mobile mesenchymal cells. This process is called the Epithelial-to-Mesenchymal Transition (EMT). Once released, they migrate along heavily stereotyped pathways throughout the entire embryo to seed distant organs.
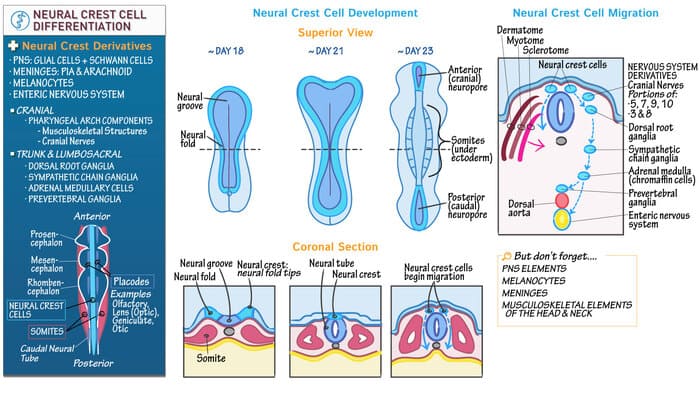
4.2 Neural Crest Derivatives
The sheer variety of tissues generated by the neural crest is staggering. To master this for exams, utilize the MAGIC COPS mnemonic, or categorize them systematically:
- Schwann cells: The myelinating cells of the PNS (Unlike oligodendrocytes which myelinate the CNS).
- All Ganglia: Dorsal Root Ganglia (DRG) containing pseudounipolar sensory neurons, Autonomic ganglia (Sympathetic chain, parasympathetic ganglia), and Enteric ganglia (Auerbach's and Meissner's plexuses in the gut).
- Adrenal Medulla: Chromaffin cells that secrete epinephrine/norepinephrine are technically modified post-ganglionic sympathetic neurons derived from the crest.
- Parafollicular (C) cells of the Thyroid: Secrete Calcitonin to lower blood calcium.
- Enterochromaffin cells: Present in the gut.
- Melanocytes: Pigment-producing cells in the skin and iris.
- Odontoblasts: Cells that produce the dentin of teeth.
- Pharyngeal Arch Cartilages: Tracheal and laryngeal cartilages, and bones of the face/skull (viscerocranium).
- Aorticopulmonary Septum: The spiral septum that divides the truncus arteriosus into the Aorta and Pulmonary Trunk. (Neural crest defects here cause severe congenital heart defects like Tetralogy of Fallot or Truncus Arteriosus).
- Leptomeninges: The inner two layers wrapping the brain: the Pia mater and Arachnoid mater. (Note: The tough outer Dura mater is derived from mesoderm).
SECTION 5: HIGH-YIELD CLINICAL EMBRYOLOGY
When the delicate timing of neural development fails to develop as intended, the resulting congenital malformations are often severe. This section covers the most heavily tested clinical pathologies.
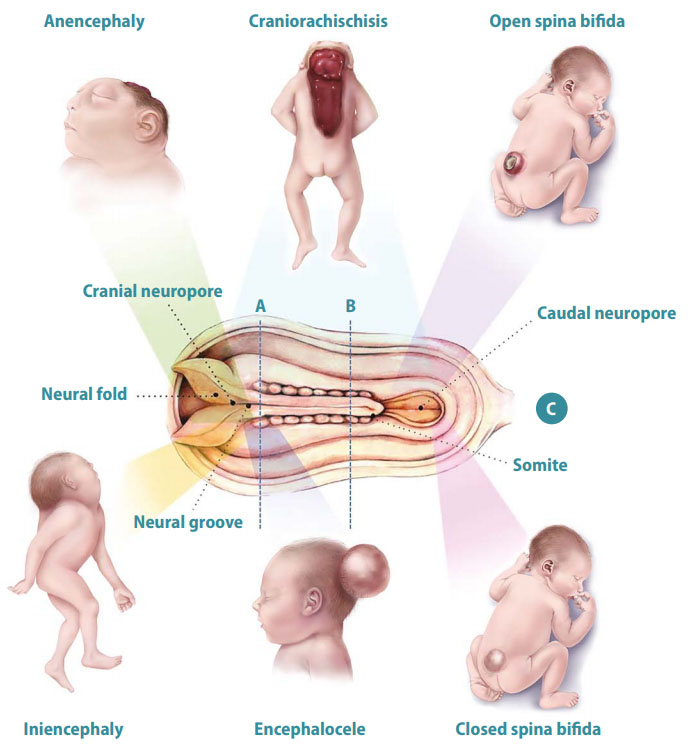
5.1 Neural Tube Defects (NTDs) & Diagnostic Markers
NTDs occur when the neural tube fails to close completely during the fourth week of gestation. The timing and location of the failure determine the specific defect and clinical severity. Maternal Folic Acid (Vitamin B9) deficiency during early pregnancy is the most significant preventable risk factor, as folate is required for nucleotide synthesis and DNA methylation during rapid cellular division.
- Spina Bifida Occulta: The mildest form. Failure of the posterior vertebral arches to fuse. The spinal cord and meninges remain in their normal position. The overlying skin is intact. It is usually asymptomatic and incidentally discovered. Clinical Sign: A tuft of hair, dimple, or birthmark over the lower lumbar spine.
- Spina Bifida Cystica: A more severe defect where tissue herniates through the bony defect, forming a cystic sac on the back.
- Meningocele: Only the meninges (dura/arachnoid) and CSF herniate into the sac. The spinal cord remains inside the vertebral canal. Neurological deficits may be absent or mild.
- Myelomeningocele: Both the meninges AND the spinal cord/nerve roots herniate into the sac. This causes severe, irreversible neurological deficits below the level of the lesion (paralysis, bowel/bladder incontinence). It is almost always associated with Chiari II malformation.
- Anencephaly: Failure of the rostral (cranial) neuropore to close. The brain and calvarium fail to develop. The fetus has a "frog-like" appearance. It is incompatible with life. Presents with maternal polyhydramnios because the fetus lacks the swallowing centers in the brain to swallow amniotic fluid.
- Encephalocele: Herniation of brain tissue and meninges through a skull defect (usually occipital).
- Elevated Alpha-Fetoprotein (AFP): In open NTDs (Myelomeningocele, Anencephaly), fetal CSF leaks into the amniotic fluid and crosses into the maternal blood.
Exception: AFP is NORMAL in Spina Bifida Occulta because the skin is intact, preventing leakage. - Elevated Acetylcholinesterase (AChE): Found in amniotic fluid. This is highly specific for neural tissue exposure and is used to confirm an open NTD if AFP is elevated.
5.2 Holoprosencephaly (Forebrain Cleavage Defect)
A failure of the embryonic prosencephalon (forebrain) to cleave into two distinct cerebral hemispheres. This represents a spectrum of structural abnormalities, from complete fusion (alobar) to partial fusion (lobar).
- Etiology: Mutations in the Sonic Hedgehog (SHH) signaling pathway. Environmental factors (maternal alcohol use, retinoic acid). Strongly associated with Patau syndrome (Trisomy 13).
- Clinical Presentation: Severe facial anomalies strictly corresponding to the severity of brain fusion. Includes cyclopia (one eye), proboscis, cleft lip/palate, and a single fused central incisor. Often fatal.
5.3 Posterior Fossa Malformations
These malformations involve defects in the development of the cerebellum and the spaces enclosing the 4th ventricle.
| Condition | Pathophysiology & Morphological Description | Associated Features |
|---|---|---|
| Dandy-Walker Malformation | Agenesis (complete absence) or severe hypoplasia of the cerebellar vermis. This leads to a massive cystic dilation of the 4th ventricle that fills the enlarged posterior fossa. | Non-communicating hydrocephalus. Presents with a noticeably enlarged skull (macrocephaly) and a bulging occiput. |
| Chiari I Malformation | Downward herniation of the cerebellar tonsils (>5mm) through the foramen magnum. Usually asymptomatic in childhood. | Often presents in young adulthood with occipital headaches exacerbated by coughing/straining. Strongly associated with Syringomyelia (fluid-filled cyst in the spinal cord). |
| Chiari II Malformation | More severe. Downward herniation of both the cerebellar vermis and tonsils, PLUS the medulla, through the foramen magnum. This physically blocks the aqueduct/4th ventricle. | Presents early in life with non-communicating hydrocephalus. Almost universally associated with Lumbar Myelomeningocele (paralysis below the defect). |
5.4 Migration Defects: Lissencephaly & Heterotopia
As discussed in Section 3, neurons must migrate outward along radial glia. When this fails, the cortex does not form properly.
- Lissencephaly ("Smooth Brain"): Failure of neuronal migration during weeks 12-24 of gestation. The brain completely lacks sulci and gyri, appearing perfectly smooth. Caused by mutations in cytoskeleton/migration proteins like Doublecortin (DCX) or Reelin. Results in severe intellectual disability, failure to thrive, and intractable seizures.
- Periventricular Nodular Heterotopia: A milder migration defect. Clusters of neurons fail to migrate and remain stranded deep in the brain, right next to the ventricles. These ectopic clusters of gray matter look like nodules on an MRI. Because they are excitable neurons in the wrong place, they frequently cause epilepsy/seizures, though intelligence may be normal.
SECTION 6: CONGENITAL HYDROCEPHALUS
Hydrocephalus is the abnormal accumulation of Cerebrospinal Fluid (CSF) within the ventricular system, leading to increased intracranial pressure (ICP). In infants (whose cranial sutures have not yet fused), this causes the head to rapidly expand.
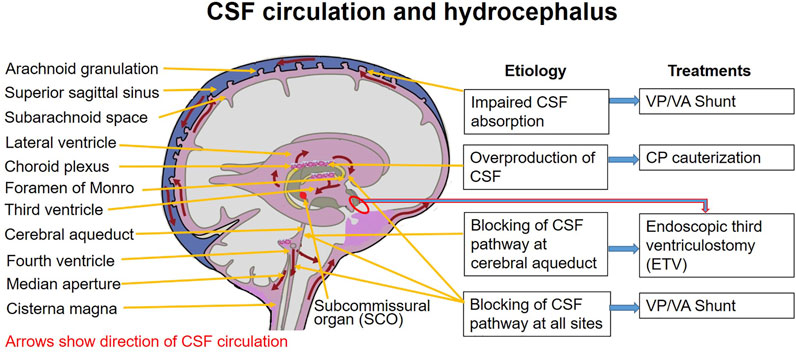
6.1 CSF Flow Pathway
To understand hydrocephalus, one must perfectly memorize the normal flow of CSF:
- Produced continuously by the Choroid Plexus primarily in the Lateral Ventricles.
- Passes through the Interventricular Foramen of Monro into the Third Ventricle.
- Flows through the narrow Cerebral Aqueduct of Sylvius (in the midbrain) into the Fourth Ventricle.
- Exits the Fourth Ventricle via three openings: the two lateral Foramina of Luschka and the single midline Foramen of Magendie.
- Enters the Subarachnoid Space, bathing the brain and spinal cord.
- Absorbed back into the venous bloodstream via the Arachnoid Granulations projecting into the Superior Sagittal Sinus.
6.2 Types of Hydrocephalus
Hydrocephalus is strictly categorized based on exactly where the blockage occurs relative to the ventricular exit doors (Luschka/Magendie).
The blockage occurs within the ventricular system itself. The CSF cannot communicate with the subarachnoid space.
- Aqueductal Stenosis: The most common cause of congenital hydrocephalus. The cerebral aqueduct narrows or is blocked. Result: Lateral and 3rd ventricles balloon massively, while the 4th ventricle remains completely normal size.
- Chiari II Malformation: Herniated cerebellar tissue physically crushes the 4th ventricle and aqueduct.
- Dandy-Walker: The cystic 4th ventricle blocks normal outflow.
The entire ventricular system is open and communicates freely. The problem is a failure of absorption at the end of the line, or massive overproduction.
- Arachnoid Granulation Failure: Scarring from prior meningitis (e.g., pneumococcal or TB) or subarachnoid hemorrhage physically clogs the arachnoid villi, preventing CSF from entering the venous blood. ALL ventricles enlarge symmetrically.
- Choroid Plexus Papilloma: A benign tumor that secretes massive amounts of CSF, overwhelming the absorption capacity.
INTEGRATION POINTS TO NOTE
- INTEGRATION POINT 1: FOLATE & METHYLATION
Why is Folate (Vitamin B9) so important for preventing Neural Tube Defects? Rapid cellular division requires vast amounts of nucleotides for DNA replication. Folate provides the single-carbon methyl groups required for the synthesis of Thymidine. Without folate, DNA replication halts, and the fast-growing neural folds cannot physically bridge the gap to fuse. - INTEGRATION POINT 2: VENTRICLE DILATION TELLS YOU WHERE THE OBSTRUCTION IS
If a clinical vignette describes massive lateral and third ventricles, but a totally normal or small fourth ventricle, the blockage MUST be exactly between them—at the Cerebral Aqueduct of Sylvius. - INTEGRATION POINT 3: CHIARI II vs. CHIARI I
Chiari I is mostly asymptomatic until adulthood and involves only the tonsils. Chiari II is a pediatric emergency involving the vermis, tonsils, and medulla, presenting with hydrocephalus and is inextricably linked to Myelomeningocele.
MNEMONICS SUMMARY
- SAME DAVE: Sensory Afferent, Motor Efferent. Dorsal Afferent, Ventral Efferent.
- 3 P's, 1 M, 1 R (3 to 5 Vesicles): Prosencephalon -> Telencephalon, Diencephalon. Mesencephalon -> Mesencephalon. Rhombencephalon -> Metencephalon, Myelencephalon.
- Tell Di, Mes Met Myel: The 5 secondary vesicles in order from top to bottom.
- MAGIC COPS (Neural Crest Derivatives): Melanocytes, Aorticopulmonary septum, Ganglia, Iris (pigment), Chromaffin cells, Cranial nerves, Odontoblasts, Parafollicular C cells, Schwann cells.
COMPARISON TABLES
| Microglia vs. Macroglia | Chiari I vs. Chiari II |
|---|---|
|
Microglia: Mesodermal origin. CNS Macrophages. Phagocytosis. Macroglia: Neuroectodermal origin. Includes Astrocytes (BBB, repair) and Oligodendrocytes (Myelin in CNS). |
Chiari I: Tonsils only (>5mm). Adult onset. Headaches. Associated with Syringomyelia. Chiari II: Vermis + Tonsils + Medulla. Infant onset. Hydrocephalus. Associated with Myelomeningocele. |
CLINICAL CORRELATIONS BY EMBRYOLOGICAL TIMING
- Week 3: Gastrulation occurs (formation of 3 germ layers). The Notochord induces the neural plate.
- Week 4: Neural tube begins closing. Primary brain vesicles form. (Critical window for NTDs; folate deficiency strikes here).
- Week 5: Secondary brain vesicles form. Neural crest cell migration is highly active.
- Weeks 12-24: Intense neuronal migration along radial glia. (Critical window for Lissencephaly).
REVIEW CHECKLIST
Before the exam, ensure you can:
- Trace the sensory dorsal horn back to its molecular inducer (BMP) and its embryonic structure (Alar plate).
- Trace the motor ventral horn back to its molecular inducer (SHH) and its embryonic structure (Basal plate).
- Draw the flow of CSF from the lateral ventricles down to the superior sagittal sinus.
- Explain the exact pathophysiology of why maternal folate deficiency causes anencephaly or myelomeningocele.
- Distinguish between the 3 primary vesicles and the 5 secondary vesicles, naming every adult derivative for each.
List of References
- Sadler, T. W. (2018). Langman's Medical Embryology (14th ed.). Wolters Kluwer. (Key resource for neurulation, neural crest migration, and SHH pathway specifics).
- Schoenwolf, G. C., Bleyl, S. B., Brauer, P. R., & Francis-West, P. H. (2020). Larsen's Human Embryology (6th ed.). Elsevier. (Reference for primary and secondary vesiculation and ventricular mapping).
- Haines, D. E. (2018). Neuroanatomy in Clinical Context: An Atlas of Structures, Sections, Systems, and Syndromes (10th ed.). Wolters Kluwer. (Reference for adult derivatives of the embryonic brain vesicles).
- Moore, K. L., Persaud, T. V. N., & Torchia, M. G. (2019). The Developing Human: Clinically Oriented Embryology (11th ed.). Elsevier. (Reference for congenital anomalies like Holoprosencephaly and Hirschsprung disease).