Digestion and Absorption of Dietary Carbohydrates
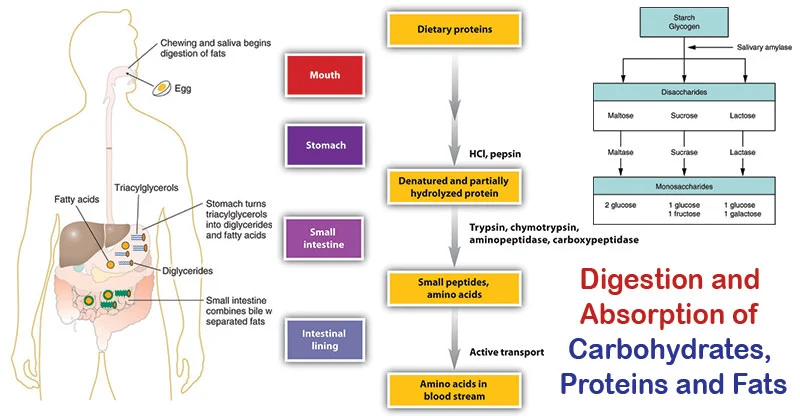
Before carbohydrates can be used by the body, complex forms (polysaccharides and disaccharides) must be broken down into monosaccharides for absorption. This process begins in the mouth and continues in the small intestine.
1. Digestion
Digestion involves the enzymatic hydrolysis of glycosidic bonds.
-
In the Mouth:
- Mechanical digestion (chewing).
- Salivary alpha-amylase (Ptyalin): Begins the breakdown of starch into smaller polysaccharides (dextrins) and some maltose. It's inactivated by stomach acid.
- In the Stomach: No significant carbohydrate digestion occurs here due to the acidic environment.
-
In the Small Intestine: This is where the bulk of carbohydrate digestion takes place.
- Pancreatic alpha-amylaseme: Continues breaking down starch and dextrins into maltose and other small polymers.
- Brush Border Enzymes: Located on the microvilli of intestinal cells, these are responsible for the final breakdown into monosaccharides.
- Maltase: Hydrolyzes maltose → two glucose molecules.
- Sucrase: Hydrolyzes sucrose → one glucose and one fructose.
- Lactase: Hydrolyzes lactose → one glucose and one galactose.
- Alpha-dextrinase (Isomaltase): Hydrolyzes alpha-1,6 bonds in limit dextrins, releasing glucose.
The end products are almost exclusively monosaccharides: glucose, fructose, and galactose.
2. Absorption
Monosaccharides are absorbed by intestinal epithelial cells (enterocytes) and then transported into the bloodstream.
-
Glucose and Galactose Absorption:
- Primarily absorbed by secondary active transport via the SGLT1 (Sodium-Glucose Cotransporter 1) protein. This requires energy and co-transports Na⁺ ions.
- From the enterocyte, they exit into the bloodstream via facilitated diffusion through the GLUT2 transporter.
-
Fructose Absorption:
- Absorbed solely by facilitated diffusion via the GLUT5 transporter. This does not require energy.
- From the enterocyte, it also exits into the bloodstream via the GLUT2 transporter.
3. Transport to the Liver
Once absorbed, these monosaccharides enter the hepatic portal vein, which carries them directly to the liver. The liver is the primary site for fructose and galactose metabolism, converting them into glucose or its intermediates.
Clinical Significance:
- Lactose Intolerance: A deficiency of the enzyme lactase, leading to maldigestion of lactose.
- Pancreatic Insufficiency: Conditions like cystic fibrosis can lead to maldigestion of starch.
- SGLT1 Deficiency: A rare genetic disorder where glucose and galactose cannot be absorbed.
Fates of Absorbed Monosaccharides (Especially Glucose)
After absorption, our monosaccharides (primarily glucose) enter the bloodstream. The body then has several crucial "fates" or pathways for this glucose, depending on energy needs and hormonal signals.
Visualizing the "Fates":
Imagine glucose as a central hub. From this hub, it can be directed down different "roads":
- Road 1: "Burn it for immediate power!" (Glycolysis → TCA → ETC)
- Road 2: "Store it for a quick pick-me-up!" (Glycogenesis)
- Road 3: "Pack it away for a rainy day!" (Conversion to Fat)
- Road 4: "Build other essential parts!" (Pentose Phosphate Pathway)
Energy Production (Oxidation):
- Goal: To generate ATP.
- Pathways: Glycolysis → Pyruvate Oxidation → TCA Cycle → Oxidative Phosphorylation.
- When: Continuously in most cells, especially during high energy demand.
Storage as Glycogen (Glycogenesis):
- Goal: To store excess glucose for later use.
- Where: Primarily in the liver and skeletal muscles.
- When: When blood glucose is high (e.g., after a meal), stimulated by insulin.
Conversion to Fat (Lipogenesis):
- Goal: To store excess energy in a long-term form when glycogen stores are full.
- Pathways: Glucose is converted to Acetyl-CoA, which is then used for fatty acid synthesis and stored as triglycerides in adipose tissue.
- When: When carbohydrate intake consistently exceeds energy needs.
Formation of Other Biomolecules (e.g., via Pentose Phosphate Pathway):
- Goal: To provide precursors for other essential molecules.
- Pathway: Pentose Phosphate Pathway (PPP) / Hexose Monophosphate Shunt (HMP Shunt):
- Produces NADPH: Crucial for biosynthesis (e.g., fatty acids) and protecting cells from oxidative stress.
- Produces Ribose-5-phosphate: A key component of nucleotides (DNA, RNA) and coenzymes (ATP, NADH).
- When: Continuously in cells with high demand for NADPH (e.g., liver, adipose tissue) or nucleotide synthesis.
Click Here To play the Game of Glycolysis
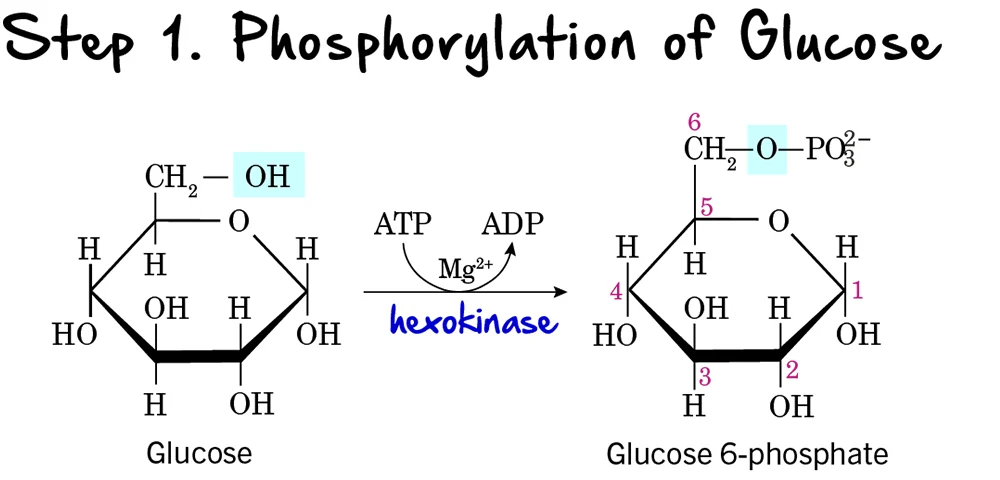
Glycolysis: Step 1 - Phosphorylation of Glucose
The first step in glycolysis is a crucial preparatory reaction, where glucose is activated and trapped within the cell.
Reaction:
Glucose is phosphorylated on its carbon 6 (C6) hydroxyl group to form Glucose 6-phosphate (G6P). This reaction consumes one molecule of ATP.
Key Features of Step 1:
-
Enzyme: The phosphorylation is catalyzed by kinases, which are enzymes that transfer a phosphate group from ATP.
- Hexokinase: Found in most tissues. It has a high affinity for glucose, meaning it can efficiently phosphorylate glucose even at low concentrations. It is inhibited by its product, glucose-6-phosphate.
-
Glucokinase: Primarily found in the liver and pancreatic beta cells. It has a lower affinity for glucose, acting only when blood glucose levels are high. It is not inhibited by glucose-6-phosphate, allowing the liver to continue taking up glucose. Both enzymes require
Mg²⁺as a cofactor.
- Intermediate Formed: Glucose 6-phosphate
- ATP Change: -1 ATP (One ATP molecule is consumed). This is the first "investment" in the energy-investment phase.
Purpose of Phosphorylation:
- Traps Glucose in the Cell: The addition of a negatively charged phosphate group prevents glucose 6-phosphate from easily crossing the cell membrane. Once phosphorylated, glucose is effectively "locked" inside the cell.
- Activates Glucose: The phosphate group makes glucose more reactive and unstable, priming it for subsequent enzymatic reactions.
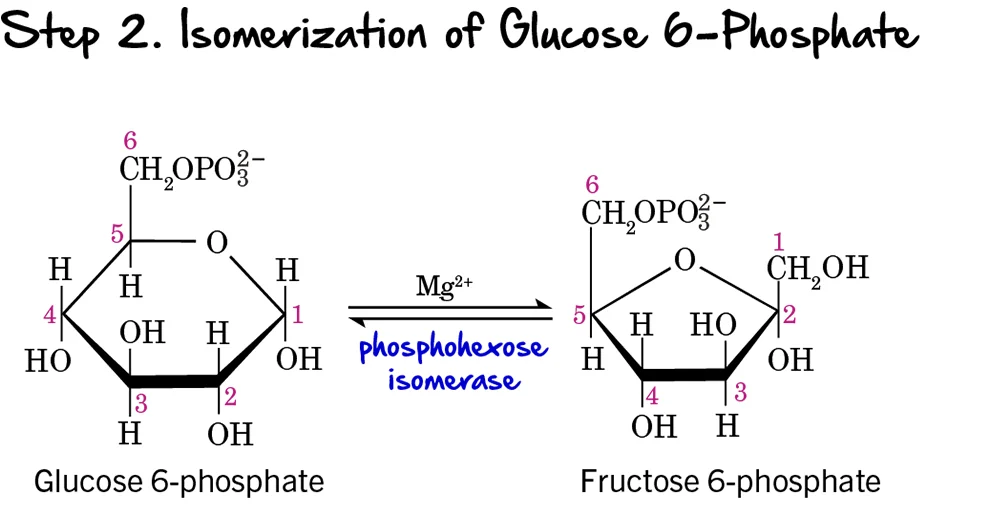
Step 2 - Isomerization of Glucose 6-Phosphate
Following the phosphorylation of glucose, the next step involves a rearrangement of the molecule from an aldose to a ketose.
Reaction:
Glucose 6-phosphate (an aldose) is isomerized (rearranged) into Fructose 6-phosphate (a ketose). This reaction is reversible.
Key Features of Step 2:
-
Enzyme: The reaction is catalyzed by Phosphohexose Isomerase (also known as Phosphoglucose Isomerase or PGI). It requires
Mg²⁺as a cofactor. - Intermediate Formed: Fructose 6-phosphate
- ATP Change: 0 ATP (No ATP is consumed or produced in this step).
Purpose of Isomerization:
This isomerization is crucial because it sets up the molecule for the next two steps in glycolysis:
- It creates a primary alcohol group at carbon 1 (C1) of fructose 6-phosphate, which can then be phosphorylated in the next step.
- It prepares the molecule for symmetric cleavage in a later step (Step 4), allowing it to be split into two 3-carbon units. If glucose 6-phosphate were cleaved directly, it would result in unequal 2-carbon and 4-carbon fragments.
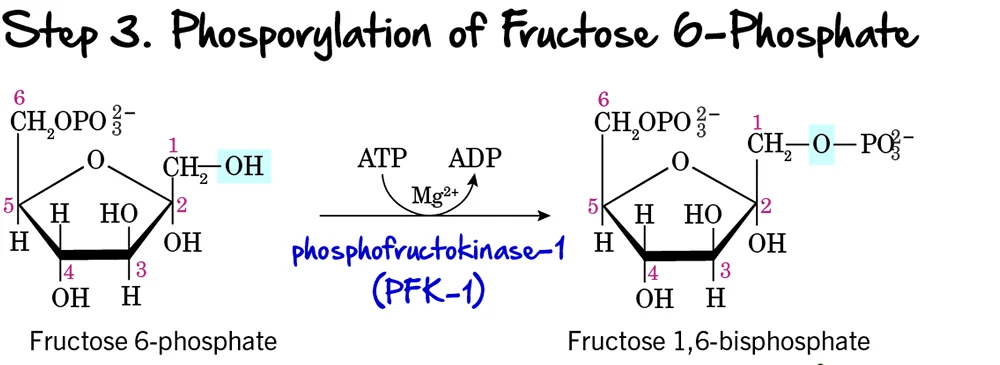
Step 3 - Phosphorylation of Fructose 6-Phosphate
This is a critical and highly regulated step in glycolysis, often considered the "committed step" of the pathway.
Reaction:
Fructose 6-phosphate undergoes a second phosphorylation, this time at its carbon 1 (C1) hydroxyl group, to form Fructose 1,6-bisphosphate. This reaction consumes another molecule of ATP.
Key Features of Step 3:
-
Enzyme: The enzyme catalyzing this reaction is Phosphofructokinase-1 (PFK-1). This is a crucial enzyme and a major regulatory point. It requires ATP as the phosphate donor and
Mg²⁺as a cofactor. - Intermediate Formed: Fructose 1,6-bisphosphate
- ATP Change: -1 ATP (Another ATP molecule is invested, bringing the total to 2 ATP).
Purpose of this Step:
- Commitment to Glycolysis: The formation of Fructose 1,6-bisphosphate is the committed step. Once formed, this molecule is generally destined to proceed through the rest of the glycolytic pathway.
- Preparation for Cleavage: Having phosphate groups on both ends (C1 and C6) is essential for the symmetrical cleavage that occurs in the next step.
Regulation of PFK-1:
PFK-1 is a key control point because its activity dictates the overall rate of glycolysis. It is allosterically regulated:
-
Activation:
- High AMP (Adenosine Monophosphate): Indicates low cellular energy, so PFK-1 is activated to increase ATP production.
- Fructose 2,6-bisphosphate: A potent allosteric activator, signaling high levels of available glucose.
-
Inhibition:
- High ATP: Indicates ample cellular energy. ATP binds to an allosteric site on PFK-1, reducing its activity.
- High Citrate: An intermediate of the Krebs Cycle, indicating that energy precursors are abundant, thus signaling to slow glycolysis.
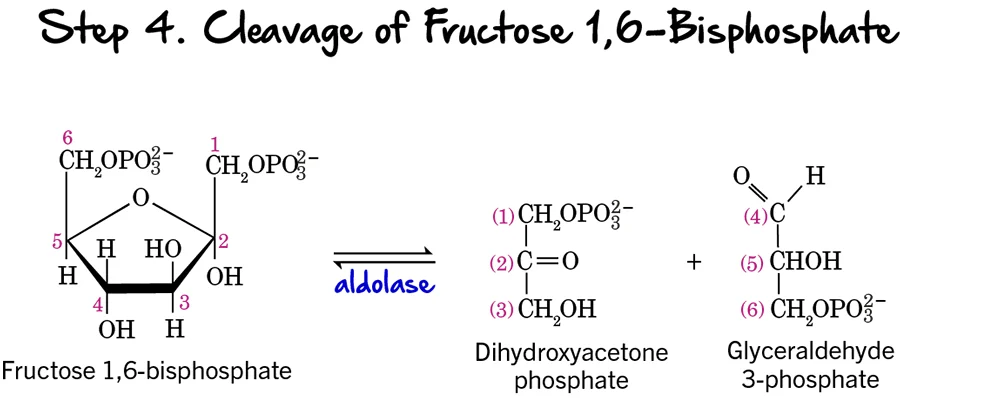
Step 4 - Cleavage of Fructose 1,6-bisphosphate
After two phosphorylation steps and an isomerization, the 6-carbon sugar is now ready to be split into two 3-carbon molecules, marking the true "lysis" of glycolysis.
Reaction:
Fructose 1,6-bisphosphate (a 6-carbon sugar) is cleaved into two distinct 3-carbon phosphorylated sugars:
- Glyceraldehyde 3-phosphate (GAP), an aldose sugar.
- Dihydroxyacetone phosphate (DHAP), a ketose sugar.
Key Features of Step 4:
- Enzyme: The enzyme catalyzing this reversible cleavage is Aldolase. The name refers to its ability to catalyze an aldol cleavage reaction.
- Intermediates Formed: Glyceraldehyde 3-phosphate (GAP) and Dihydroxyacetone phosphate (DHAP).
- ATP Change: 0 ATP (No ATP is consumed or produced).
Purpose of the Cleavage:
This step is crucial because it takes the single 6-carbon sugar and converts it into two 3-carbon molecules. These two molecules will then proceed through the second, "energy payoff" stage. The previous isomerization to fructose 6-phosphate (Step 2) was essential to enable this symmetrical cleavage into two triose phosphates, making the rest of the pathway more efficient.
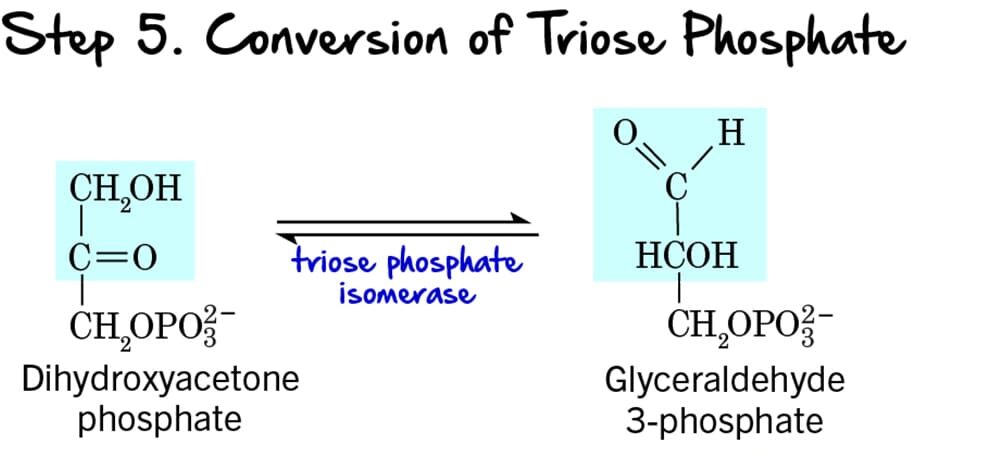
Step 5 - Interconversion of Triose Phosphates
Following the cleavage of Fructose 1,6-bisphosphate (Step 4), two different 3-carbon sugars are produced: Dihydroxyacetone phosphate (DHAP) and Glyceraldehyde 3-phosphate (GAP). However, only GAP can directly proceed into the next steps of glycolysis. This step ensures that both molecules can be utilized.
Reaction:
Dihydroxyacetone phosphate (DHAP), a ketose, is reversibly isomerized into Glyceraldehyde 3-phosphate (GAP), an aldose.
Key Features of Step 5:
- Enzyme: The enzyme catalyzing this reversible isomerization is Triose Phosphate Isomerase (TPI). This enzyme is remarkably efficient, catalyzing the reaction at a rate close to the diffusion limit.
- Intermediate Formed: Through this reaction, all the carbon atoms from the initial glucose molecule are now in the form of Glyceraldehyde 3-phosphate (GAP). From one glucose molecule, we now have two molecules of GAP ready to enter the energy payoff phase.
- ATP Change: 0 ATP (No ATP is consumed or produced).
Purpose of the Interconversion:
This isomerization is crucial because:
- Ensures Efficient Pathway Progression: Only Glyceraldehyde 3-phosphate can move forward. By converting DHAP to GAP, the cell ensures that all carbon atoms from the original glucose are processed efficiently.
- Maintains Balance: The reaction is reversible, maintaining an equilibrium between DHAP and GAP, although the subsequent rapid consumption of GAP drives the equilibrium towards GAP formation.
End of Energy Investment Phase
This concludes the Energy Investment Phase (Reactions 1-5). We have now invested 2 ATP and converted one 6-carbon glucose into two 3-carbon Glyceraldehyde 3-phosphate molecules. The pathway is now ready to enter the Energy Payoff Phase.
The Energy Payoff Phase:
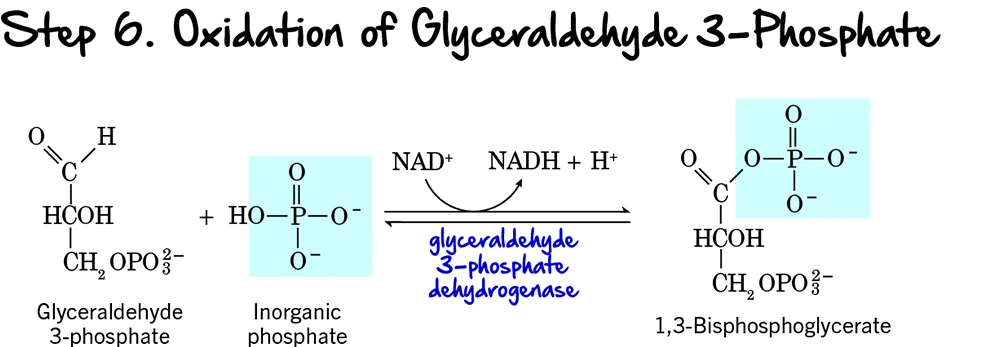
Step 6 (Oxidation and Phosphorylation of Glyceraldehyde 3-Phosphate)
We're now entering the Energy Payoff Phase of glycolysis! This is where the cell starts to recover its ATP investment and generate reducing power. Step 6 is the first reaction in this phase, and it's a crucial one as it involves both an oxidation event and the formation of a high-energy phosphate compound.
Reaction:
Each molecule of Glyceraldehyde 3-phosphate (GAP) undergoes a two-part transformation:
- Oxidation: The aldehyde group of GAP is oxidized to a carboxyl group.
- Phosphorylation: An inorganic phosphate (Pᵢ) group (not from ATP) is added to this newly formed carboxyl group, creating an acyl phosphate bond.
Key Features of Step 6:
- Enzyme: The enzyme catalyzing this reaction is Glyceraldehyde 3-phosphate Dehydrogenase.
- Intermediate Formed: 1,3-Bisphosphoglycerate (1,3-BPG).
- ATP Change: 0 ATP directly.
- NADH Production: +1 NADH is produced per molecule of GAP. Since each glucose yields two GAP molecules, this step generates a total of 2 NADH per glucose.
Purpose of this Step:
- Generation of Reducing Power (NADH): This is the only redox reaction in glycolysis. The electrons released during the oxidation of GAP are captured by NAD⁺, forming NADH. NADH is a crucial electron carrier that will later produce ATP in the electron transport chain (under aerobic conditions).
- Formation of a High-Energy Phosphate Bond: The newly formed bond at carbon 1 of 1,3-BPG is an acyl phosphate bond. This is a high-energy bond, meaning its hydrolysis releases significant free energy, which will be harnessed in the next step to synthesize ATP.
- Primer for ATP Synthesis: By creating 1,3-BPG with its high-energy phosphate, this step sets the stage for the first ATP generation in glycolysis via substrate-level phosphorylation.
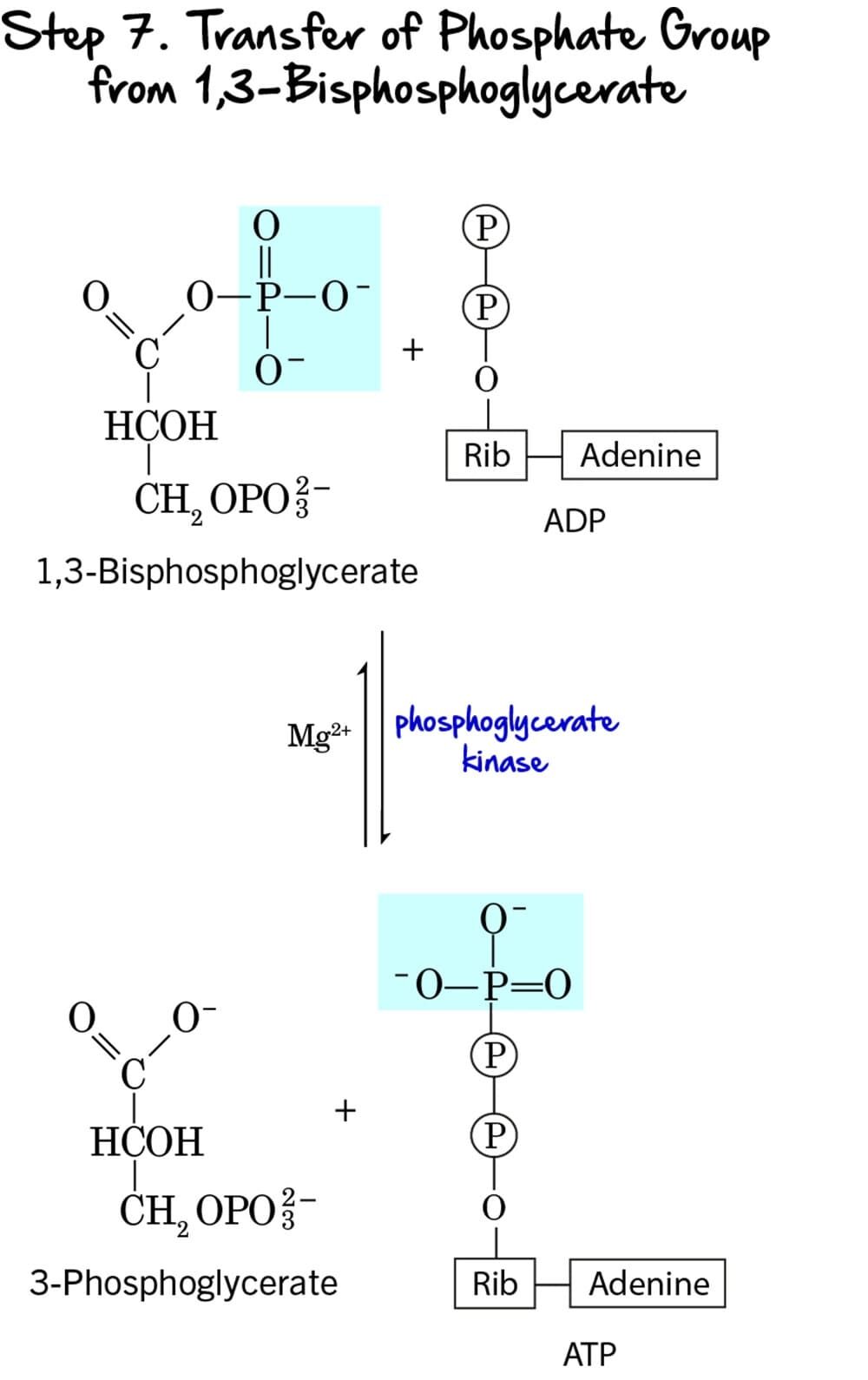
Step 7 - Substrate-Level Phosphorylation
This step marks the first direct production of ATP in glycolysis, utilizing the high-energy phosphate bond generated in the previous step.
Reaction:
The high-energy phosphate group from the C1 position of 1,3-Bisphosphoglycerate (1,3-BPG) is transferred to ADP, forming ATP. The remaining molecule is 3-Phosphoglycerate.
Key Features of Step 7:
-
Enzyme: The reaction is catalyzed by Phosphoglycerate Kinase. It requires
Mg²⁺as a cofactor. - Intermediate Formed: 3-Phosphoglycerate.
- ATP Change: +1 ATP is generated per molecule of 1,3-BPG. Since two molecules of 1,3-BPG are produced from each glucose, this step generates a total of 2 ATP per glucose.
- Mechanism: This is a classic example of substrate-level phosphorylation. ATP is formed directly from the transfer of a high-energy phosphate group from a substrate (1,3-BPG) to ADP.
Purpose of this Step:
- ATP Generation: This is the first actual ATP gain in glycolysis, partially recovering the energy invested in the preparatory phase.
- Energy Recovery: The energy released from the hydrolysis of the high-energy acyl phosphate bond in 1,3-BPG is efficiently captured to synthesize ATP.
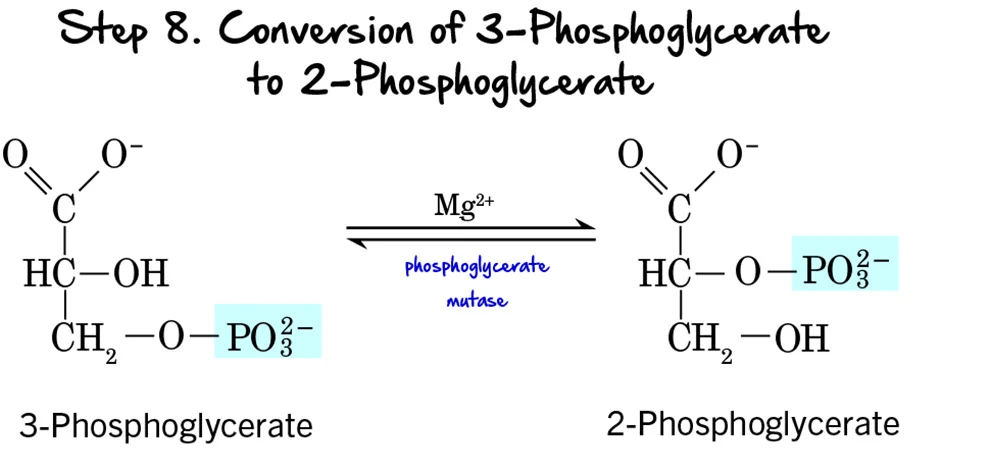
Step 8 - Migration of the Phosphate Group
After the first ATP-generating step, the molecule undergoes a structural rearrangement of its phosphate group to prepare for the next high-energy bond formation.
Reaction:
The phosphate group on 3-Phosphoglycerate moves from the carbon at position 3 to the carbon at position 2, forming 2-Phosphoglycerate. This is an intramolecular rearrangement.
Key Features of Step 8:
-
Enzyme: The reaction is catalyzed by Phosphoglycerate Mutase. Mutases are a class of isomerases that catalyze the transfer of a functional group within the same molecule. This enzyme requires
Mg²⁺as a cofactor. - Intermediate Formed: 2-Phosphoglycerate.
- ATP Change: 0 ATP (No ATP is consumed or produced).
Purpose of the Phosphate Migration:
This rearrangement is crucial for the subsequent steps:
- Positions for Dehydration: Moving the phosphate group to the C2 position places it in a strategic location to allow for the formation of a high-energy phosphate bond in the next step. It creates the necessary conditions for the dehydration reaction that follows.
- Increased Energy Potential: While 2-phosphoglycerate itself doesn't contain a high-energy bond, its structure is primed to become one through the elimination of water.
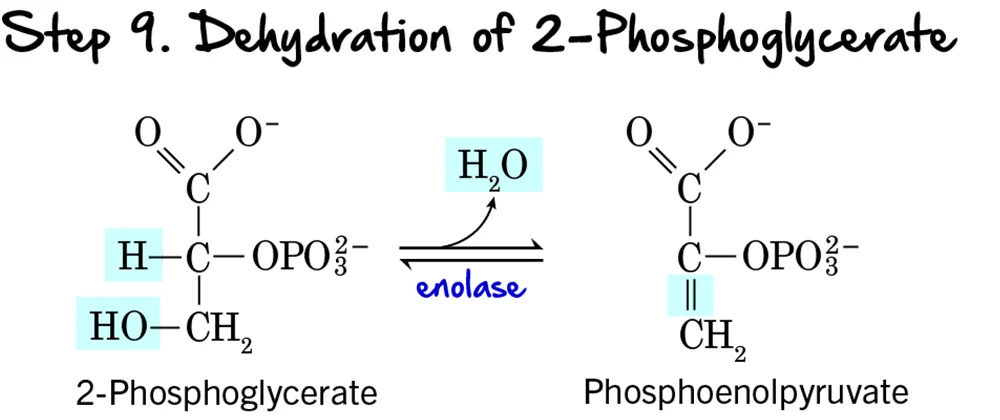
Step 9 - Dehydration of 2-Phosphoglycerate
Following the migration of the phosphate group, the molecule undergoes a dehydration reaction, which significantly raises the phosphoryl transfer potential of the phosphate group.
Reaction:
A molecule of water (H₂O) is removed from 2-Phosphoglycerate. This dehydration reaction creates a double bond within the molecule and forms the high-energy compound Phosphoenolpyruvate (PEP), which contains an "enol phosphate" bond.
Key Features of Step 9:
- Enzyme: The reaction is catalyzed by Enolase.
- Intermediate Formed: Phosphoenolpyruvate (PEP).
- ATP Change: 0 ATP (No ATP is consumed or produced).
Purpose of the Dehydration:
- Creation of a High-Energy Phosphate Bond: This is the most important outcome. The removal of water redistributes energy within the molecule, transforming a low-energy phosphate bond into a high-energy enol phosphate bond. The ΔG°' for the hydrolysis of PEP's phosphate bond is one of the highest in biochemistry.
- Preparation for ATP Synthesis: By forming PEP, the molecule is now poised to donate its phosphate group to ADP to generate ATP in the final step of glycolysis.
Clinical Relevance:
Enolase is inhibited by fluoride ions. This property is exploited in clinical settings: when blood samples are collected for glucose measurement, fluoride is often added to the collection tube to prevent glycolysis by red blood cells, ensuring the measured glucose concentration is accurate.
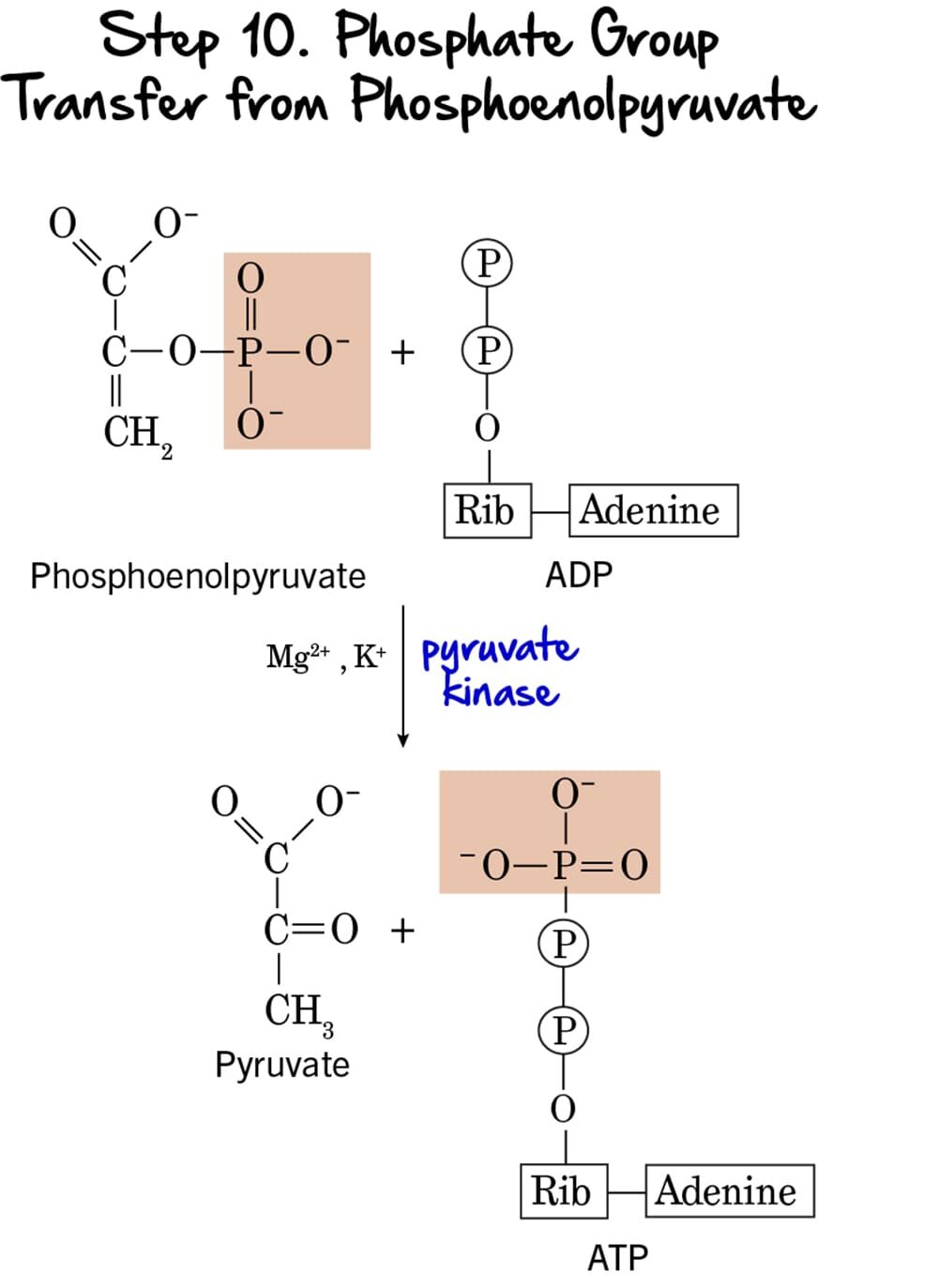
Step 10 - Phosphate Group Transfer from Phosphoenolpyruvate
This is the second and final ATP-generating step in glycolysis, again utilizing substrate-level phosphorylation to produce ATP and the ultimate end-product, pyruvate.
Reaction:
The high-energy phosphate group from Phosphoenolpyruvate (PEP) is transferred to ADP, yielding another molecule of ATP. The product remaining is Pyruvate. The initial enol form of pyruvate immediately tautomerizes to the more stable keto form.
Key Features of Step 10:
-
Enzyme: The reaction is catalyzed by Pyruvate Kinase, another key regulatory enzyme. It requires
Mg²⁺andK⁺as cofactors. - Intermediate Formed: Pyruvate.
- ATP Change: +1 ATP is generated per molecule of PEP. Since two molecules of PEP are produced from each glucose, this step generates a total of 2 ATP per glucose.
- Mechanism: This is the second instance of substrate-level phosphorylation in glycolysis.
Purpose of this Step:
- ATP Generation: This step provides the second net gain of ATP, completing the energy payoff.
- Formation of Pyruvate: Pyruvate is the end-product of glycolysis. Its fate depends on oxygen availability. Under aerobic conditions, it enters the mitochondria; under anaerobic conditions, it can be fermented.
- Irreversible Step & Regulation: This reaction is essentially irreversible, making Pyruvate Kinase a crucial regulatory enzyme.
Regulation of Pyruvate Kinase:
Pyruvate kinase is tightly regulated to control the flow of carbon through glycolysis:
-
Activation:
- Fructose 1,6-bisphosphate: This is a classic example of feed-forward activation. The product of PFK-1 (an earlier step) activates pyruvate kinase, ensuring intermediates are quickly processed.
-
Inhibition:
- High ATP: Signals abundant energy.
- Acetyl-CoA: An indicator of high energy status.
- Long-chain fatty acids: Another alternative fuel source.
Summary of Glycolysis (Net Reaction per glucose):
→
2 Pyruvate + 2 NADH + 2 H⁺ + 2 ATP
Glycolysis has now broken down one 6-carbon glucose molecule into two 3-carbon pyruvate molecules, produced a net of 2 ATP molecules, and generated 2 NADH molecules for further energy production.
Biochemistry: Glycolysis Exam
Test your knowledge with these 40 questions.
Glycolysis Exam
Question 1/40
Exam Complete!
Here are your results, .
Your Score
38/40
95%